Evolution
| Part of the Biology series on |
| Evolution |
| Mechanisms and processes |
|---|
|
Adaptation |
| Research and history |
|
Introduction |
| Evolutionary biology fields |
|
Cladistics |
| Biology portal · |
Evolution (also known as biological, genetic or organic evolution)[1][2] is the change in the inherited traits of a population of organisms through successive generations.[3] This change results from interactions between processes which introduce variation into a population, and other processes which remove it. As a result, variants with particular traits become more, or less, common. A trait is a particular characteristic, anatomical, biochemical or behavioural, that is the result of gene–environment interaction.
The main source of variation is mutation, which changes the base pair sequences of genes. These altered genes can be passed on through reproduction, and give rise to alternative varieties, or alleles, of traits in organisms. Another source of variation is genetic recombination which shuffles the genes into new combinations that can result in organisms exhibiting different traits. Under certain circumstances, variation can also be increased by the transfer of genes between species,[4][5] and by the extremely rare, but significant, wholesale incorporation of genomes through endosymbiosis.[6][7]
Two main processes cause variants to become more common or rare in a population. One is natural selection, through which traits that aid survival and reproduction become more common, while traits that hinder survival and reproduction become more rare. Natural selection occurs because only a small proportion of individuals in each generation will survive and reproduce, since resources are limited and organisms produce many more offspring than their environment can support. Over many generations, heritable variation in traits is filtered by natural selection and the beneficial changes are successively retained through differential survival and reproduction. This iterative process adjusts traits so they become better suited to an organism's environment: these adjustments are called adaptations.[8]
However, not all change is adaptive. Another cause of evolution is genetic drift, which leads to random changes in how common traits are in a population. Genetic drift is most important when traits do not strongly influence survival, particularly so in small populations where chance plays a disproportionate role in the frequency of traits passed on to the next generation.[9][10] Genetic drift is important in the neutral theory of molecular evolution, and plays a role in the molecular clocks that are used in phylogenetic studies.
A key process in evolution is speciation, in which a single ancestral species splits and diversifies into multiple new species, and there are several modes through which this occurs. Ultimately, all living (and extinct) species are descended from a common ancestor via a long series of speciation events. These events stretch back in a diverse "tree of life" that has grown over the 3.5 billion years in which life has existed on Earth.[11][12][13][14] This is visible in anatomical, genetic and other similarities between groups of organisms, geographical distribution of related species, the fossil record and the recorded genetic changes in living organisms over many generations.
Evolutionary biologists document the fact that evolution occurs, and also develop and test theories that explain its causes. The study of evolutionary biology began in the mid-nineteenth century, when research into the fossil record and the diversity of living organisms convinced most scientists that species changed over time.[15][16] The mechanism driving these changes remained unclear until the theory of natural selection was independently proposed by Charles Darwin and Alfred Wallace. In 1859, Darwin's seminal work On the Origin of Species brought the new theory of evolution by natural selection to a wide audience,[17] leading to the overwhelming acceptance of evolution among scientists.[18][19][20][21] In the 1930s, Darwinian natural selection became understood in combination with Mendelian inheritance, forming the modern evolutionary synthesis,[22] which connected the substrate of evolution (inherited genetics) and the mechanism of evolution (natural selection). This powerful explanatory and predictive theory has become the central organizing principle of modern biology, directing research and providing a unifying explanation for the history and diversity of life on Earth.[19][20][23] Evolution is applied and studied in fields as diverse as agriculture, anthropology, conservation biology, ecology, medicine, paleontology, philosophy, and psychology along with other specific topics in the previous listed fields.
Contents |
History of evolutionary thought

The roots of scientific thinking can be dated to at least the 6th century BCE, with the Greek philosopher Anaximander.[24] However, the growth of modern biology out of natural history is fairly recent. As biological knowledge grew in the 18th century, proto-evolutionary ideas were set out by a few natural philosophers such as Pierre Maupertuis in 1745 and Erasmus Darwin in 1796.[25] The ideas of the biologist Jean-Baptiste Lamarck about transmutation of species influenced radicals, but were rejected by mainstream scientists. Charles Darwin formulated his idea of natural selection in 1838 and was still developing his theory in 1858 when Alfred Russel Wallace sent him a similar theory, and both were presented to the Linnean Society of London in separate papers.[26] At the end of 1859, Darwin's publication of On the Origin of Species explained natural selection in detail and presented evidence leading to increasingly wide acceptance of the occurrence of evolution.
Debate about the mechanisms of evolution continued, and Darwin could not explain the source of the heritable variations which would be acted on by natural selection. Like Lamarck, he thought that parents passed on adaptations acquired during their lifetimes,[27] a theory which was subsequently dubbed Lamarckism.[28] In the 1880s, August Weismann's experiments indicated that changes from use and disuse were not heritable, and Lamarckism gradually fell from favour.[29][30] More significantly, Darwin could not account for how traits were passed down from generation to generation. In 1865 Gregor Mendel found that traits were inherited in a predictable manner.[31] When Mendel's work was rediscovered in 1900s, disagreements over the rate of evolution predicted by early geneticists and biometricians led to a rift between the Mendelian and Darwinian models of evolution.
Yet it was the rediscovery of Gregor Mendel's pioneering work on the fundamentals of genetics (of which Darwin and Wallace were unaware) by Hugo de Vries and others in the early 1900s that provided the impetus for a better understanding of how variation occurs in plant and animal traits. That variation is the main fuel used by natural selection to shape the wide variety of adaptive traits observed in organic life. Even though Hugo de Vries and other early geneticists rejected gradual natural selection, their rediscovery of and subsequent work on genetics eventually provided a solid basis on which the theory of evolution stood even more convincingly than when it was originally proposed.[32]
The apparent contradiction between Darwin's theory of evolution by natural selection and Mendel's work was reconciled in the 1920s and 1930s by evolutionary biologists such as J.B.S. Haldane, Sewall Wright, and particularly Ronald Fisher, who set the foundations for the establishment of the field of population genetics. The end result was a combination of evolution by natural selection and Mendelian inheritance, the modern evolutionary synthesis.[33] In the 1940s, the identification of DNA as the genetic material by Oswald Avery and colleagues and the subsequent publication of the structure of DNA by James Watson and Francis Crick in 1953, demonstrated the physical basis for inheritance. Since then, genetics and molecular biology have become core parts of evolutionary biology and have revolutionized the field of phylogenetics.[22]
In its early history, evolutionary biology primarily drew in scientists from traditional taxonomically oriented disciplines, whose specialist training in particular organisms addressed general questions in evolution. As evolutionary biology expanded as an academic discipline, particularly after the development of the modern evolutionary synthesis, it began to draw more widely from the biological sciences.[22] Currently the study of evolutionary biology involves scientists from fields as diverse as biochemistry, ecology, genetics and physiology, and evolutionary concepts are used in even more distant disciplines such as psychology, medicine, philosophy and computer science. In the 21st century, current research in evolutionary biology deals with several areas where the modern evolutionary synthesis may need modification or extension, such as assessing the relative importance of various ideas on the unit of selection and evolvability and how to fully incorporate the findings of evolutionary developmental biology.[34][35]
Heredity

Evolution in organisms occurs through changes in heritable traits – particular characteristics of an organism. In humans, for example, eye color is an inherited characteristic and an individual might inherit the "brown-eye trait" from one of their parents.[36] Inherited traits are controlled by genes and the complete set of genes within an organism's genome is called its genotype.[37]
The complete set of observable traits that make up the structure and behavior of an organism is called its phenotype. These traits come from the interaction of its genotype with the environment.[38] As a result, many aspects of an organism's phenotype are not inherited. For example, suntanned skin comes from the interaction between a person's genotype and sunlight; thus, suntans are not passed on to people's children. However, some people tan more easily than others, due to differences in their genotype; a striking example are people with the inherited trait of albinism, who do not tan at all and are very sensitive to sunburn.[39]
Heritable traits are passed from one generation to the next via DNA, a molecule that encodes genetic information.[37] DNA is a long polymer composed of four types of bases. The sequence of bases along a particular DNA molecule specify the genetic information, in a manner similar to a sequence of letters spelling out a sentence. Before a cell divides, the DNA is copied, so that each of the resulting two cells will inherit the DNA sequence.
Portions of a DNA molecule that specify a single functional unit are called genes; different genes have different sequences of bases. Within cells, the long strands of DNA form condensed structures called chromosomes. The specific location of a DNA sequence within a chromosome is known as a locus. If the DNA sequence at a locus varies between individuals, the different forms of this sequence are called alleles. DNA sequences can change through mutations, producing new alleles. If a mutation occurs within a gene, the new allele may affect the trait that the gene controls, altering the phenotype of the organism.
However, while this simple correspondence between an allele and a trait works in some cases, most traits are more complex and are controlled by multiple interacting genes.[40][41] The study of such complex traits is a major area of current genetic research. Another unsolved question in genetics is whether or not epigenetics is important in evolution. Epigenetics is when a trait is inherited without there being any change in gene sequences.[42]
Variation
An individual organism's phenotype results from both its genotype and the influence from the environment it has lived in. A substantial part of the variation in phenotypes in a population is caused by the differences between their genotypes.[41] The modern evolutionary synthesis defines evolution as the change over time in this genetic variation. The frequency of one particular allele will fluctuate, becoming more or less prevalent relative to other forms of that gene. Evolutionary forces act by driving these changes in allele frequency in one direction or another. Variation disappears when a new allele reaches the point of fixation — when it either disappears from the population or replaces the ancestral allele entirely.[43]
Variation comes from mutations in genetic material, migration between populations (gene flow), and the reshuffling of genes through sexual reproduction. Variation also comes from exchanges of genes between different species; for example, through horizontal gene transfer in bacteria, and hybridization in plants.[44] Despite the constant introduction of variation through these processes, most of the genome of a species is identical in all individuals of that species.[45] However, even relatively small changes in genotype can lead to dramatic changes in phenotype: for example, chimpanzees and humans differ in only about 5% of their genomes.[46]
Mutation
Random mutations constantly occur in the genomes of organisms; these mutations create genetic variation. Mutations are changes in the DNA sequence of a cell's genome and are caused by radiation, viruses, transposons and mutagenic chemicals, as well as errors that occur during meiosis or DNA replication.[47][48][49] These mutations involve several different types of change in DNA sequences; these can either have no effect, alter the product of a gene, or prevent the gene from functioning. Studies in the fly Drosophila melanogaster suggest that if a mutation changes a protein produced by a gene, this will probably be harmful, with about 70 percent of these mutations having damaging effects, and the remainder being either neutral or weakly beneficial.[50] Due to the damaging effects that mutations can have on cells, organisms have evolved mechanisms such as DNA repair to remove mutations.[47] Therefore, the optimal mutation rate for a species is a trade-off between costs of a high mutation rate, such as deleterious mutations, and the metabolic costs of maintaining systems to reduce the mutation rate, such as DNA repair enzymes.[51] Viruses that use RNA as their genetic material have rapid mutation rates,[52] which can be an advantage since these viruses will evolve constantly and rapidly, and thus evade the defensive responses of e.g. the human immune system.[53]
Mutations can involve large sections of a chromosome becoming duplicated (usually by genetic recombination), which can introduce extra copies of a gene into a genome.[54] Extra copies of genes are a major source of the raw material needed for new genes to evolve.[55] This is important because most new genes evolve within gene families from pre-existing genes that share common ancestors.[56] For example, the human eye uses four genes to make structures that sense light: three for color vision and one for night vision; all four are descended from a single ancestral gene.[57] New genes can be created from an ancestral gene when a duplicate copy mutates and acquires a new function. This process is easier once a gene has been duplicated because it increases the redundancy of the system; one gene in the pair can acquire a new function while the other copy continues to perform its original function.[58][59] Other types of mutation can even create entirely new genes from previously noncoding DNA.[60][61] The creation of new genes can also involve small parts of several genes being duplicated, with these fragments then recombining to form new combinations with new functions.[62][63] When new genes are assembled from shuffling pre-existing parts, domains act as modules with simple independent functions, which can be mixed together creating new combinations with new and complex functions.[64] For example, polyketide synthases are large enzymes that make antibiotics; they contain up to one hundred independent domains that each catalyze one step in the overall process, like a step in an assembly line.[65]
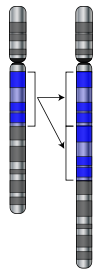
Changes in chromosome number may involve even larger mutations, where segments of the DNA within chromosomes break and then rearrange. For example, two chromosomes in the Homo genus fused to produce human chromosome 2; this fusion did not occur in the lineage of the other apes, and they retain these separate chromosomes.[66] In evolution, the most important role of such chromosomal rearrangements may be to accelerate the divergence of a population into new species by making populations less likely to interbreed, and thereby preserving genetic differences between these populations.[67]
Sequences of DNA that can move about the genome, such as transposons, make up a major fraction of the genetic material of plants and animals, and may have been important in the evolution of genomes.[68] For example, more than a million copies of the Alu sequence are present in the human genome, and these sequences have now been recruited to perform functions such as regulating gene expression.[69] Another effect of these mobile DNA sequences is that when they move within a genome, they can mutate or delete existing genes and thereby produce genetic diversity.[48]
Sex and recombination
In asexual organisms, genes are inherited together, or linked, as they cannot mix with genes of other organisms during reproduction. In contrast, the offspring of sexual organisms contain random mixtures of their parents' chromosomes that are produced through independent assortment. In a related process called homologous recombination, sexual organisms exchange DNA between two matching chromosomes.[70] Recombination and reassortment do not alter allele frequencies, but instead change which alleles are associated with each other, producing offspring with new combinations of alleles.[71] Sex usually increases genetic variation and may increase the rate of evolution.[72][73] However, asexuality is advantageous in some environments as it can evolve in previously sexual animals.[74] Here, asexuality might allow the two sets of alleles in their genome to diverge and gain different functions.[75]
Recombination allows even alleles that are close together in a strand of DNA to be inherited independently. However, the rate of recombination is low (approximately two events per chromosome per generation). As a result, genes close together on a chromosome may not always be shuffled away from each other, and genes that are close together tend to be inherited together, a phenomenon known as linkage.[76] This tendency is measured by finding how often two alleles occur together on a single chromosome, which is called their linkage disequilibrium. A set of alleles that is usually inherited in a group is called a haplotype. This can be important when one allele in a particular haplotype is strongly beneficial: natural selection can drive a selective sweep that will also cause the other alleles in the haplotype to become more common in the population; this effect is called genetic hitchhiking.[77]
When alleles cannot be separated by recombination – such as in mammalian Y chromosomes, which pass intact from fathers to sons – harmful mutations accumulate.[78][79] By breaking up allele combinations, sexual reproduction allows the removal of harmful mutations and the retention of beneficial mutations.[80] In addition, recombination and reassortment can produce individuals with new and advantageous gene combinations. These positive effects are balanced by the fact that sex reduces an organism's reproductive rate, can cause mutations and may separate beneficial combinations of genes.[80] The reasons for the evolution of sexual reproduction are therefore unclear and this question is still an active area of research in evolutionary biology,[81][82] that has prompted ideas such as the Red Queen hypothesis.[83]
Population genetics


From a genetic viewpoint, evolution is a generation-to-generation change in the frequencies of alleles within a population that shares a common gene pool.[84] A population is a localized group of individuals belonging to the same species. For example, all of the moths of the same species living in an isolated forest represent a population. A single gene in this population may have several alternate forms, which account for variations between the phenotypes of the organisms. An example might be a gene for coloration in moths that has two alleles: black and white. A gene pool is the complete set of alleles for a gene in a single population; the allele frequency measures the fraction of the gene pool composed of a single allele (for example, what fraction of moth coloration genes are the black allele). Evolution occurs when there are changes in the frequencies of alleles within a population of interbreeding organisms; for example, the allele for black color in a population of moths becoming more common.
To understand the mechanisms that cause a population to evolve, it is useful to consider what conditions are required for a population not to evolve. The Hardy-Weinberg principle states that the frequencies of alleles (variations in a gene) in a sufficiently large population will remain constant if the only forces acting on that population are the random reshuffling of alleles during the formation of the sperm or egg, and the random combination of the alleles in these sex cells during fertilization.[85] Such a population is said to be in Hardy-Weinberg equilibrium; it is not evolving.[86]
Gene flow

Gene flow is the exchange of genes between populations, which are usually of the same species.[88] Examples of gene flow within a species include the migration and then breeding of organisms, or the exchange of pollen. Gene transfer between species includes the formation of hybrid organisms and horizontal gene transfer.
Migration into or out of a population can change allele frequencies, as well as introducing genetic variation into a population. Immigration may add new genetic material to the established gene pool of a population. Conversely, emigration may remove genetic material. As barriers to reproduction between two diverging populations are required for the populations to become new species, gene flow may slow this process by spreading genetic differences between the populations. Gene flow is hindered by mountain ranges, oceans and deserts or even man-made structures such as the Great Wall of China, which has hindered the flow of plant genes.[89]
Depending on how far two species have diverged since their most recent common ancestor, it may still be possible for them to produce offspring, as with horses and donkeys mating to produce mules.[90] Such hybrids are generally infertile, due to the two different sets of chromosomes being unable to pair up during meiosis. In this case, closely related species may regularly interbreed, but hybrids will be selected against and the species will remain distinct. However, viable hybrids are occasionally formed and these new species can either have properties intermediate between their parent species, or possess a totally new phenotype.[91] The importance of hybridization in creating new species of animals is unclear, although cases have been seen in many types of animals,[92] with the gray tree frog being a particularly well-studied example.[93]
Hybridization is, however, an important means of speciation in plants, since polyploidy (having more than two copies of each chromosome) is tolerated in plants more readily than in animals.[94][95] Polyploidy is important in hybrids as it allows reproduction, with the two different sets of chromosomes each being able to pair with an identical partner during meiosis.[96] Polyploids also have more genetic diversity, which allows them to avoid inbreeding depression in small populations.[97]
Horizontal gene transfer is the transfer of genetic material from one organism to another organism that is not its offspring; this is most common among bacteria.[98] In medicine, this contributes to the spread of antibiotic resistance, as when one bacteria acquires resistance genes it can rapidly transfer them to other species.[1] Horizontal transfer of genes from bacteria to eukaryotes such as the yeast Saccharomyces cerevisiae and the adzuki bean beetle Callosobruchus chinensis may also have occurred.[99][100] An example of larger-scale transfers are the eukaryotic bdelloid rotifers, which appear to have received a range of genes from bacteria, fungi, and plants.[101] Viruses can also carry DNA between organisms, allowing transfer of genes even across biological domains.[102] Large-scale gene transfer has also occurred between the ancestors of eukaryotic cells and prokaryotes, during the acquisition of chloroplasts and mitochondria.[103]
Mechanisms
The two main mechanisms that produce evolution are natural selection and genetic drift. Natural selection is the process which favors genes that aid survival and reproduction. Genetic drift is the random change in the frequency of alleles, caused by the random sampling of a generation's genes during reproduction. The relative importance of natural selection and genetic drift in a population varies depending on the strength of the selection and the effective population size, which is the number of individuals capable of breeding.[104] Natural selection usually predominates in large populations, whereas genetic drift dominates in small populations. The dominance of genetic drift in small populations can even lead to the fixation of slightly deleterious mutations.[105] As a result, changing population size can dramatically influence the course of evolution. Population bottlenecks, where the population shrinks temporarily and therefore loses genetic variation, result in a more uniform population.[43]
Natural selection

Natural selection is the process by which genetic mutations that enhance reproduction become, and remain, more common in successive generations of a population. It has often been called a "self-evident" mechanism because it necessarily follows from three simple facts:
- Heritable variation exists within populations of organisms.
- Organisms produce more offspring than can survive.
- These offspring vary in their ability to survive and reproduce.
These conditions produce competition between organisms for survival and reproduction. Consequently, organisms with traits that give them an advantage over their competitors pass these advantageous traits on, while traits that do not confer an advantage are not passed on to the next generation.[106]
The central concept of natural selection is the evolutionary fitness of an organism.[107] Fitness is measured by an organism's ability to survive and reproduce, which determines the size of its genetic contribution to the next generation.[107] However, fitness is not the same as the total number of offspring: instead fitness is indicated by the proportion of subsequent generations that carry an organism's genes.[108] For example, if an organism could survive well and reproduce rapidly, but its offspring were all too small and weak to survive, this organism would make little genetic contribution to future generations and would thus have low fitness.[107]
If an allele increases fitness more than the other alleles of that gene, then with each generation this allele will become more common within the population. These traits are said to be "selected for". Examples of traits that can increase fitness are enhanced survival, and increased fecundity. Conversely, the lower fitness caused by having a less beneficial or deleterious allele results in this allele becoming rarer — they are "selected against".[109] Importantly, the fitness of an allele is not a fixed characteristic; if the environment changes, previously neutral or harmful traits may become beneficial and previously beneficial traits become harmful.[3] However, even if the direction of selection does reverse in this way, traits that were lost in the past may not re-evolve in an identical form (see Dollo's law).[110][111]

Natural selection within a population for a trait that can vary across a range of values, such as height, can be categorized into three different types. The first is directional selection, which is a shift in the average value of a trait over time — for example organisms slowly getting taller.[112] Secondly, disruptive selection is selection for extreme trait values and often results in two different values becoming most common, with selection against the average value. This would be when either short or tall organisms had an advantage, but not those of medium height. Finally, in stabilizing selection there is selection against extreme trait values on both ends, which causes a decrease in variance around the average value and less diversity.[106][113] This would, for example, cause organisms to slowly become all the same height.
A special case of natural selection is sexual selection, which is selection for any trait that increases mating success by increasing the attractiveness of an organism to potential mates.[114] Traits that evolved through sexual selection are particularly prominent in males of some animal species, despite traits such as cumbersome antlers, mating calls or bright colors that attract predators, decreasing the survival of individual males.[115] This survival disadvantage is balanced by higher reproductive success in males that show these hard to fake, sexually selected traits.[116]
Natural selection most generally makes nature the measure against which individuals, and individual traits, are more or less likely to survive. "Nature" in this sense refers to an ecosystem, that is, a system in which organisms interact with every other element, physical as well as biological, in their local environment. Eugene Odum, a founder of ecology, defined an ecosystem as: "Any unit that includes all of the organisms...in a given area interacting with the physical environment so that a flow of energy leads to clearly defined trophic structure, biotic diversity, and material cycles (ie: exchange of materials between living and nonliving parts) within the system."[117] Each population within an ecosystem occupies a distinct niche, or position, with distinct relationships to other parts of the system. These relationships involve the life history of the organism, its position in the food chain, and its geographic range. This broad understanding of nature enables scientists to delineate specific forces which, together, comprise natural selection.
An active area of research is the unit of selection, with natural selection being proposed to work at the level of genes, cells, individual organisms, groups of organisms and species.[118][119] None of these are mutually exclusive and selection can act on multiple levels simultaneously.[120] An example of selection occurring below the level of the individual organism are genes called transposons, which can replicate and spread throughout a genome.[121] Selection at a level above the individual, such as group selection, may allow the evolution of co-operation, as discussed below.[122]
Genetic drift

Genetic drift is the change in allele frequency from one generation to the next that occurs because alleles in offspring are a random sample of those in the parents, as well as from the role that chance plays in determining whether a given individual will survive and reproduce. In mathematical terms, alleles are subject to sampling error. As a result, when selective forces are absent or relatively weak, allele frequencies tend to "drift" upward or downward randomly (in a random walk). This drift halts when an allele eventually becomes fixed, either by disappearing from the population, or replacing the other alleles entirely. Genetic drift may therefore eliminate some alleles from a population due to chance alone. Even in the absence of selective forces, genetic drift can cause two separate populations that began with the same genetic structure to drift apart into two divergent populations with different sets of alleles.[123]
The time for an allele to become fixed by genetic drift depends on population size, with fixation occurring more rapidly in smaller populations.[124] The precise measure of population that is important is called the effective population size. The effective population is always smaller than the total population since it takes into account factors such as the level of inbreeding, the number of animals that are too old or young to breed, and the lower probability of animals that live far apart managing to mate with each other.[125]
An example when genetic drift is probably of central importance in determining a trait is the loss of pigments from animals that live in caves, a change that produces no obvious advantage or disadvantage in complete darkness.[126] However, it is usually difficult to measure the relative importance of selection and drift,[127] so the comparative importance of these two forces in driving evolutionary change is an area of current research.[128] These investigations were prompted by the neutral theory of molecular evolution, which proposed that most evolutionary changes are the result of the fixation of neutral mutations that do not have any immediate effects on the fitness of an organism.[129] Hence, in this model, most genetic changes in a population are the result of constant mutation pressure and genetic drift.[130] This form of the neutral theory is now largely abandoned, since it does not seem to fit the genetic variation seen in nature.[131][132] However, a more recent and better-supported version of this model is the nearly neutral theory, where most mutations only have small effects on fitness.[106]
Outcomes
Evolution influences every aspect of the form and behavior of organisms. Most prominent are the specific behavioral and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding predators or attracting mates. Organisms can also respond to selection by co-operating with each other, usually by aiding their relatives or engaging in mutually beneficial symbiosis. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed.
These outcomes of evolution are sometimes divided into macroevolution, which is evolution that occurs at or above the level of species, such as extinction and speciation, and microevolution, which is smaller evolutionary changes, such as adaptations, within a species or population.[133] In general, macroevolution is regarded as the outcome of long periods of microevolution.[134] Thus, the distinction between micro- and macroevolution is not a fundamental one – the difference is simply the time involved.[135] However, in macroevolution, the traits of the entire species may be important. For instance, a large amount of variation among individuals allows a species to rapidly adapt to new habitats, lessening the chance of it going extinct, while a wide geographic range increases the chance of speciation, by making it more likely that part of the population will become isolated. In this sense, microevolution and macroevolution might involve selection at different levels – with microevolution acting on genes and organisms, versus macroevolutionary processes such as species selection acting on entire species and affecting their rates of speciation and extinction.[136][137][138]
A common misconception is that evolution has goals or long-term plans; realistically however, evolution has no long-term goal and does not necessarily produce greater complexity.[139][140] Although complex species have evolved, they occur as a side effect of the overall number of organisms increasing, and simple forms of life still remain more common in the biosphere.[141] For example, the overwhelming majority of species are microscopic prokaryotes, which form about half the world's biomass despite their small size,[142] and constitute the vast majority of Earth's biodiversity.[143] Simple organisms have therefore been the dominant form of life on Earth throughout its history and continue to be the main form of life up to the present day, with complex life only appearing more diverse because it is more noticeable.[144] Indeed, the evolution of microorganisms is particularly important to modern evolutionary research, since their rapid reproduction allows the study of experimental evolution and the observation of evolution and adaptation in real time.[145][146]
Adaptation
Adaptation is one of the basic phenomena of biology,[147] and is the process whereby an organism becomes better suited to its habitat.[148][149] Also, the term adaptation may refer to a trait that is important for an organism's survival. For example, the adaptation of horses' teeth to the grinding of grass, or the ability of horses to run fast and escape predators. By using the term adaptation for the evolutionary process, and adaptive trait for the product (the bodily part or function), the two senses of the word may be distinguished. Adaptations are produced by natural selection.[150] The following definitions are due to Theodosius Dobzhansky.
- 1. Adaptation is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats.[151]
- 2. Adaptedness is the state of being adapted: the degree to which an organism is able to live and reproduce in a given set of habitats.[152]
- 3. An adaptive trait is an aspect of the developmental pattern of the organism which enables or enhances the probability of that organism surviving and reproducing.[153]
Adaptation may cause either the gain of a new feature, or the loss of an ancestral feature. An example that shows both types of change is bacterial adaptation to antibiotic selection, with genetic changes causing antibiotic resistance by both modifying the target of the drug, or increasing the activity of transporters that pump the drug out of the cell.[154] Other striking examples are the bacteria Escherichia coli evolving the ability to use citric acid as a nutrient in a long-term laboratory experiment,[155] Flavobacterium evolving a novel enzyme that allows these bacteria to grow on the by-products of nylon manufacturing,[156][157] and the soil bacterium Sphingobium evolving an entirely new metabolic pathway that degrades the synthetic pesticide pentachlorophenol.[158][159] An interesting but still controversial idea is that some adaptations might increase the ability of organisms to generate genetic diversity and adapt by natural selection (increasing organisms' evolvability).[160][161]

Adaptation occurs through the gradual modification of existing structures. Consequently, structures with similar internal organization may have different functions in related organisms. This is the result of a single ancestral structure being adapted to function in different ways. The bones within bat wings, for example, are very similar to those in mice feet and primate hands, due to the descent of all these structures from a common mammalian ancestor.[163] However, since all living organisms are related to some extent,[164] even organs that appear to have little or no structural similarity, such as arthropod, squid and vertebrate eyes, or the limbs and wings of arthropods and vertebrates, can depend on a common set of homologous genes that control their assembly and function; this is called deep homology.[165][166]
During adaptation, some structures may lose their original function and become vestigial structures.[167] Such structures may have little or no function in a current species, yet have a clear function in ancestral species, or other closely related species. Examples include pseudogenes,[168] the non-functional remains of eyes in blind cave-dwelling fish,[169] wings in flightless birds,[170] and the presence of hip bones in whales and snakes.[162] Examples of vestigial structures in humans include wisdom teeth,[171] the coccyx,[167] the vermiform appendix,[167] and other behavioral vestiges such as goose bumps,[172] and primitive reflexes.[173][174][175][176]
However, many traits that appear to be simple adaptations are in fact exaptations: structures originally adapted for one function, but which coincidentally became somewhat useful for some other function in the process.[177] One example is the African lizard Holaspis guentheri, which developed an extremely flat head for hiding in crevices, as can be seen by looking at its near relatives. However, in this species, the head has become so flattened that it assists in gliding from tree to tree—an exaptation.[177] Within cells, molecular machines such as the bacterial flagella[178] and protein sorting machinery[179] evolved by the recruitment of several pre-existing proteins that previously had different functions.[133] Another example is the recruitment of enzymes from glycolysis and xenobiotic metabolism to serve as structural proteins called crystallins within the lenses of organisms' eyes.[180][181]
A critical principle of ecology is that of competitive exclusion: no two species can occupy the same niche in the same environment for a long time.[182] Consequently, natural selection will tend to force species to adapt to different ecological niches. This may mean that, for example, two species of cichlid fish adapt to live in different habitats, which will minimize the competition between them for food.[183]
An area of current investigation in evolutionary developmental biology is the developmental basis of adaptations and exaptations.[184] This research addresses the origin and evolution of embryonic development and how modifications of development and developmental processes produce novel features.[185] These studies have shown that evolution can alter development to create new structures, such as embryonic bone structures that develop into the jaw in other animals instead forming part of the middle ear in mammals.[186] It is also possible for structures that have been lost in evolution to reappear due to changes in developmental genes, such as a mutation in chickens causing embryos to grow teeth similar to those of crocodiles.[187] It is now becoming clear that most alterations in the form of organisms are due to changes in a small set of conserved genes.[188]
Co-evolution

Interactions between organisms can produce both conflict and co-operation. When the interaction is between pairs of species, such as a pathogen and a host, or a predator and its prey, these species can develop matched sets of adaptations. Here, the evolution of one species causes adaptations in a second species. These changes in the second species then, in turn, cause new adaptations in the first species. This cycle of selection and response is called co-evolution.[189] An example is the production of tetrodotoxin in the rough-skinned newt and the evolution of tetrodotoxin resistance in its predator, the common garter snake. In this predator-prey pair, an evolutionary arms race has produced high levels of toxin in the newt and correspondingly high levels of toxin resistance in the snake.[190]
Co-operation
However, not all interactions between species involve conflict.[191] Many cases of mutually beneficial interactions have evolved. For instance, an extreme cooperation exists between plants and the mycorrhizal fungi that grow on their roots and aid the plant in absorbing nutrients from the soil.[192] This is a reciprocal relationship as the plants provide the fungi with sugars from photosynthesis. Here, the fungi actually grow inside plant cells, allowing them to exchange nutrients with their hosts, while sending signals that suppress the plant immune system.[193]
Coalitions between organisms of the same species have also evolved. An extreme case is the eusociality found in social insects, such as bees, termites and ants, where sterile insects feed and guard the small number of organisms in a colony that are able to reproduce. On an even smaller scale, the somatic cells that make up the body of an animal limit their reproduction so they can maintain a stable organism, which then supports a small number of the animal's germ cells to produce offspring. Here, somatic cells respond to specific signals that instruct them whether to grow, remain as they are, or die. If cells ignore these signals and multiply inappropriately, their uncontrolled growth causes cancer.[47]
Such cooperation within species may have evolved through the process of kin selection, which is where one organism acts to help raise a relative's offspring.[194] This activity is selected for because if the helping individual contains alleles which promote the helping activity, it is likely that its kin will also contain these alleles and thus those alleles will be passed on.[195] Other processes that may promote cooperation include group selection, where cooperation provides benefits to a group of organisms.[196]
Speciation

Speciation is the process where a species diverges into two or more descendant species.[197] Evolutionary biologists view species as statistical phenomena and not categories or types. This view is counterintuitive since the classical idea of species is still widely held, with a species seen as a class of organisms exemplified by a "type specimen" that bears all the traits common to this species. Instead, a species is now defined as a separately evolving lineage that forms a single gene pool. Although properties such as genetics and morphology are used to help separate closely related lineages, this definition has fuzzy boundaries.[198] Indeed, the exact definition of the term "species" is still controversial, particularly in prokaryotes,[199] and this is called the species problem.[200] Biologists have proposed a range of more precise definitions, but the definition used is a pragmatic choice that depends on the particularities of the species concerned.[200] Typically the actual focus on biological study is the population, an observable interacting group of organisms, rather than a species, an observable similar group of individuals.
Speciation has been observed multiple times under both controlled laboratory conditions and in nature.[201] In sexually reproducing organisms, speciation results from reproductive isolation followed by genealogical divergence. There are four mechanisms for speciation. The most common in animals is allopatric speciation, which occurs in populations initially isolated geographically, such as by habitat fragmentation or migration. Selection under these conditions can produce very rapid changes in the appearance and behaviour of organisms.[202][203] As selection and drift act independently on populations isolated from the rest of their species, separation may eventually produce organisms that cannot interbreed.[204]
The second mechanism of speciation is peripatric speciation, which occurs when small populations of organisms become isolated in a new environment. This differs from allopatric speciation in that the isolated populations are numerically much smaller than the parental population. Here, the founder effect causes rapid speciation through both rapid genetic drift and selection on a small gene pool.[205]
The third mechanism of speciation is parapatric speciation. This is similar to peripatric speciation in that a small population enters a new habitat, but differs in that there is no physical separation between these two populations. Instead, speciation results from the evolution of mechanisms that reduce gene flow between the two populations.[197] Generally this occurs when there has been a drastic change in the environment within the parental species' habitat. One example is the grass Anthoxanthum odoratum, which can undergo parapatric speciation in response to localized metal pollution from mines.[206] Here, plants evolve that have resistance to high levels of metals in the soil. Selection against interbreeding with the metal-sensitive parental population produced a gradual change in the flowering time of the metal-resistant plants, which eventually produced complete reproductive isolation. Selection against hybrids between the two populations may cause reinforcement, which is the evolution of traits that promote mating within a species, as well as character displacement, which is when two species become more distinct in appearance.[207]
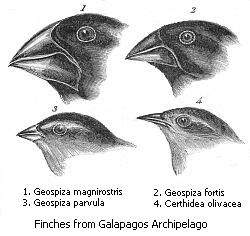
Finally, in sympatric speciation species diverge without geographic isolation or changes in habitat. This form is rare since even a small amount of gene flow may remove genetic differences between parts of a population.[208] Generally, sympatric speciation in animals requires the evolution of both genetic differences and non-random mating, to allow reproductive isolation to evolve.[209]
One type of sympatric speciation involves cross-breeding of two related species to produce a new hybrid species. This is not common in animals as animal hybrids are usually sterile. This is because during meiosis the homologous chromosomes from each parent are from different species and cannot successfully pair. However, it is more common in plants because plants often double their number of chromosomes, to form polyploids.[210] This allows the chromosomes from each parental species to form matching pairs during meiosis, since each parent's chromosomes are represented by a pair already.[211] An example of such a speciation event is when the plant species Arabidopsis thaliana and Arabidopsis arenosa cross-bred to give the new species Arabidopsis suecica.[212] This happened about 20,000 years ago,[213] and the speciation process has been repeated in the laboratory, which allows the study of the genetic mechanisms involved in this process.[214] Indeed, chromosome doubling within a species may be a common cause of reproductive isolation, as half the doubled chromosomes will be unmatched when breeding with undoubled organisms.[95]
Speciation events are important in the theory of punctuated equilibrium, which accounts for the pattern in the fossil record of short "bursts" of evolution interspersed with relatively long periods of stasis, where species remain relatively unchanged.[215] In this theory, speciation and rapid evolution are linked, with natural selection and genetic drift acting most strongly on organisms undergoing speciation in novel habitats or small populations. As a result, the periods of stasis in the fossil record correspond to the parental population, and the organisms undergoing speciation and rapid evolution are found in small populations or geographically restricted habitats, and therefore rarely being preserved as fossils.[216]
Extinction

Extinction is the disappearance of an entire species. Extinction is not an unusual event, as species regularly appear through speciation, and disappear through extinction.[217] Nearly all animal and plant species that have lived on Earth are now extinct,[218] and extinction appears to be the ultimate fate of all species.[219] These extinctions have happened continuously throughout the history of life, although the rate of extinction spikes in occasional mass extinction events.[220] The Cretaceous–Tertiary extinction event, during which the non-avian dinosaurs went extinct, is the most well-known, but the earlier Permian–Triassic extinction event was even more severe, with approximately 96 percent of species driven to extinction.[220] The Holocene extinction event is an ongoing mass extinction associated with humanity's expansion across the globe over the past few thousand years. Present-day extinction rates are 100–1000 times greater than the background rate, and up to 30 percent of species may be extinct by the mid 21st century.[221] Human activities are now the primary cause of the ongoing extinction event;[222] global warming may further accelerate it in the future.[223]
The role of extinction in evolution is not very well understood and may depend on which type of extinction is considered.[220] The causes of the continuous "low-level" extinction events, which form the majority of extinctions, may be the result of competition between species for limited resources (competitive exclusion).[22] If one species can out-compete another, this could produce species selection, with the fitter species surviving and the other species being driven to extinction.[118] The intermittent mass extinctions are also important, but instead of acting as a selective force, they drastically reduce diversity in a nonspecific manner and promote bursts of rapid evolution and speciation in survivors.[224]
Evolutionary history of life
Origin of life
The origin of life is a necessary precursor for biological evolution, but understanding that evolution occurred once organisms appeared and investigating how this happens does not depend on understanding exactly how life began.[225] The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions, but it is unclear how this occurred.[226] Not much is certain about the earliest developments in life, the structure of the first living things, or the identity and nature of any last universal common ancestor or ancestral gene pool.[227][228] Consequently, there is no scientific consensus on how life began, but proposals include self-replicating molecules such as RNA,[229] and the assembly of simple cells.[230]
Common descent

All organisms on Earth are descended from a common ancestor or ancestral gene pool.[164][231] Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events.[232] The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits, and finally, that organisms can be classified using these similarities into a hierarchy of nested groups – similar to a family tree.[17] However, modern research has suggested that, due to horizontal gene transfer, this "tree of life" may be more complicated than a simple branching tree since some genes have spread independently between distantly related species.[233][234]
Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record.[235] By comparing the anatomies of both modern and extinct species, paleontologists can infer the lineages of those species. However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.
More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and amino acids.[236] The development of molecular genetics has revealed the record of evolution left in organisms' genomes: dating when species diverged through the molecular clock produced by mutations.[237] For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 96% of their genomes and analyzing the few areas where they differ helps shed light on when the common ancestor of these species existed.[238]
Evolution of life
Despite the uncertainty on how life began, it is generally accepted that prokaryotes inhabited the Earth from approximately 3–4 billion years ago.[240][241] No obvious changes in morphology or cellular organization occurred in these organisms over the next few billion years.[242]
The eukaryotes were the next major change in cell structure. These came from ancient bacteria being engulfed by the ancestors of eukaryotic cells, in a cooperative association called endosymbiosis.[103][243] The engulfed bacteria and the host cell then underwent co-evolution, with the bacteria evolving into either mitochondria or hydrogenosomes.[244] An independent second engulfment of cyanobacterial-like organisms led to the formation of chloroplasts in algae and plants.[245] It is unknown when the first eukaryotic cells appeared though they first emerged between 1.6 – 2.7 billion years ago.
The history of life was that of the unicellular eukaryotes, prokaryotes, and archaea until about 610 million years ago when multicellular organisms began to appear in the oceans in the Ediacaran period.[240][246] The evolution of multicellularity occurred in multiple independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime moulds and myxobacteria.[247]
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over approximately 10 million years, in an event called the Cambrian explosion. Here, the majority of types of modern animals appeared in the fossil record, as well as unique lineages that subsequently became extinct.[248] Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.[249] About 500 million years ago, plants and fungi colonized the land, and were soon followed by arthropods and other animals.[250] Insects were particularly successful and even today make up the majority of animal species.[251] Amphibians first appeared around 300 million years ago, followed by early amniotes, then mammals around 200 million years ago and birds around 100 million years ago (both from "reptile"-like lineages). However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to be highly successful and dominate the Earth, with the majority of both biomass and species being prokaryotes.[143]
Applications
Evolutionary biology, and in particular the understanding of how organisms evolve through natural selection, is an area of science with many practical applications.[252] A major technological application of evolution is artificial selection, which is the intentional selection of certain traits in a population of organisms. Humans have used artificial selection for thousands of years in the domestication of plants and animals.[253] More recently, such selection has become a vital part of genetic engineering, with selectable markers such as antibiotic resistance genes being used to manipulate DNA in molecular biology. It is also possible to use repeated rounds of mutation and selection to evolve proteins with particular properties, such as modified enzymes or new antibodies, in a process called directed evolution.[254]
Understanding the changes that have occurred during organism's evolution can reveal the genes needed to construct parts of the body, genes which may be involved in human genetic disorders.[255] For example, the Mexican tetra is an albino cavefish that lost its eyesight during evolution. Breeding together different populations of this blind fish produced some offspring with functional eyes, since different mutations had occurred in the isolated populations that had evolved in different caves.[256] This helped identify genes required for vision and pigmentation, such as crystallins and the melanocortin 1 receptor.[257] Similarly, comparing the genome of the Antarctic icefish, which lacks red blood cells, to close relatives such as the Antarctic rockcod revealed genes needed to make these blood cells.[258]
As evolution can produce highly optimized processes and networks, it has many applications in computer science. Here, simulations of evolution using evolutionary algorithms and artificial life started with the work of Nils Aall Barricelli in the 1960s, and was extended by Alex Fraser, who published a series of papers on simulation of artificial selection.[259] Artificial evolution became a widely recognized optimization method as a result of the work of Ingo Rechenberg in the 1960s and early 1970s, who used evolution strategies to solve complex engineering problems.[260] Genetic algorithms in particular became popular through the writing of John Holland.[261] As academic interest grew, dramatic increases in the power of computers allowed practical applications, including the automatic evolution of computer programs.[262] Evolutionary algorithms are now used to solve multi-dimensional problems more efficiently than software produced by human designers, and also to optimize the design of systems.[263]
Social and cultural responses

In the 19th century, particularly after the publication of On the Origin of Species in 1859, the idea that life had evolved was an active source of academic debate centered on the philosophical, social and religious implications of evolution. Nowadays, the fact that organisms evolve is uncontested in the scientific literature and the modern evolutionary synthesis is widely accepted by scientists.[22] However, evolution remains a contentious concept for some theists.[265]
While various religions and denominations have reconciled their beliefs with evolution through concepts such as theistic evolution, there are creationists who believe that evolution is contradicted by the creation myths found in their respective religions and who raise various objections to evolution.[133][266][267] As had been demonstrated by responses to the publication of Vestiges of the Natural History of Creation in 1844, the most controversial aspect of evolutionary biology is the implication of human evolution that human mental and moral faculties, which had been thought purely spiritual, are not distinctly separated from those of other animals.[16] In some countries—notably the United States—these tensions between science and religion have fueled the current creation-evolution controversy, a religious conflict focusing on politics and public education.[268] While other scientific fields such as cosmology[269] and Earth science[270] also conflict with literal interpretations of many religious texts, evolutionary biology experiences significantly more opposition from religious literalists.
The teaching of evolution in American secondary school biology classes was uncommon in most of the first half of the 20th century. The Scopes Trial decision of 1925 caused the subject to become very rare in American secondary biology textbooks for a generation, but it was gradually re-introduced about a generation later and legally protected with the 1968 Epperson v. Arkansas decision. Since then, the competing religious belief of creationism was legally disallowed in secondary school curricula in various decisions in the 1970s and 1980s, but it returned in the form of intelligent design, to be excluded once again in the 2005 Kitzmiller v. Dover Area School District case.[271]
Another example somewhat associated with evolutionary theory that is now widely regarded as unwarranted is "Social Darwinism", a derogatory term associated with the 19th century Malthusian theory developed by Whig philosopher Herbert Spencer. It was later expanded by others into ideas about "survival of the fittest" in commerce and human societies as a whole, and led to claims that social inequality, sexism, racism, and imperialism were justified.[272] However, these ideas contradict Darwin's own views, and contemporary scientists and philosophers consider these ideas to be neither mandated by evolutionary theory nor supported by data.[273][274][275]
See also
References
- ↑ 1.0 1.1 Walsh T (2006). "Combinatorial genetic evolution of multiresistance". Curr. Opin. Microbiol. 9 (5): 476–82. doi:10.1016/j.mib.2006.08.009. PMID 16942901.
- ↑ Kottak, Conrad Phillip (2005). Window on Humanity: A Concise Introduction to Anthropology. The McGraw–Hill Companies, Inc, New York.
- ↑ 3.0 3.1 Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 0-87893-187-2.
- ↑ Jain, R.; Rivera, M.C.; Lake, J.A. (1999), "Horizontal gene transfer among genomes: the complexity hypothesis.", Proc Natl Acad Sci U S A 96 (7): 3801–6, doi:10.1073/pnas.96.7.3801, PMID 10097118.
- ↑ Richardson, Aaron O. and Jeffrey D. Palmer (January 2007). "Horizontal gene transfer in plants". Journal of Experimental Botany 58 (1): 1–9. doi:10.1093/jxb/erl148. PMID 17030541. http://www.sdsc.edu/~shindyal/ejc121304.pdf. Retrieved 2007-03-18.
- ↑ Margulis, Lynn (1998). The symbiotic planet: a new look at evolution. Weidenfeld & Nicolson, London.
- ↑ Sapp, J. (1994). Evolution by association: a history of symbiosis. Oxford University Press, UK.
- ↑ Ayala FJ (2007). "Darwin's greatest discovery: design without designer". Proc. Natl. Acad. Sci. U.S.A. 104 (Suppl 1): 8567–73. doi:10.1073/pnas.0701072104. PMID 17494753. PMC 1876431. http://www.pnas.org/content/104/suppl.1/8567.full.
- ↑ Evolution 101:Sampling Error and Evolution and Effects of Genetic Drift from the Understanding Evolution University of California at Berkeley
- ↑ Evolution 101: Peripatric Speciation from the Understanding Evolution webpages made by the University of California at Berkeley
- ↑ Schopf, J.W. (1999). Cradle of life: the discovery of Earth's earliest fossils.
- ↑ Woese, C. (1998). "The Universal Ancestor". PNAS 95: 6854–6859.
- ↑ Theobald, D.L. (2010). "A formal test of the theory of universal common ancestry". Nature 465: 219-222.
- ↑ Doolittle, W.F. (February, 2000). "Uprooting the tree of life". Scientific American 282: 90–95. http://shiva.msu.montana.edu/courses/mb437_537_2004_fall/docs/uprooting.pdf.
- ↑ Ian C. Johnston (1999). "History of Science: Early Modern Geology". Malaspina University-College. http://records.viu.ca/~johnstoi/darwin/sect2.htm. Retrieved 2008-01-15.
- ↑ 16.0 16.1 Bowler, Peter J. (2003). Evolution:The History of an Idea. University of California Press. ISBN 0-52023693-9.
- ↑ 17.0 17.1 Darwin, Charles (1859). On the Origin of Species (1st ed.). London: John Murray. p. 1. ISBN 0801413192. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=16.. Related earlier ideas were acknowledged in Darwin, Charles (1861). On the Origin of Species (3rd ed.). London: John Murray. xiii. ISBN 0801413192. http://darwin-online.org.uk/content/frameset?itemID=F381&viewtype=text&pageseq=20.
- ↑ AAAS Council (December 26, 1922). "AAAS Resolution: Present Scientific Status of the Theory of Evolution". American Association for the Advancement of Science. http://archives.aaas.org/docs/resolutions.php?doc_id=450.
- ↑ 19.0 19.1 "IAP Statement on the Teaching of Evolution" (PDF). The Interacademy Panel on International Issues. 2006. http://www.interacademies.net/Object.File/Master/6/150/Evolution%20statement.pdf. Retrieved 2007-04-25. Joint statement issued by the national science academies of 67 countries, including the United Kingdom's Royal Society
- ↑ 20.0 20.1 Board of Directors, American Association for the Advancement of Science (2006-02-16). "Statement on the Teaching of Evolution" (PDF). American Association for the Advancement of Science. http://www.aaas.org/news/releases/2006/pdf/0219boardstatement.pdf. from the world's largest general scientific society
- ↑ "Statements from Scientific and Scholarly Organizations". National Center for Science Education. http://ncse.com/media/voices/science.
- ↑ 22.0 22.1 22.2 22.3 22.4 Kutschera U, Niklas K (2004). "The modern theory of biological evolution: an expanded synthesis". Naturwissenschaften 91 (6): 255–76. doi:10.1007/s00114-004-0515-y. PMID 15241603.
- ↑ "Special report on evolution". New Scientist. 2008-01-19. http://www.newscientist.com/topic/evolution.
- ↑ Wright, S (1984). Evolution and the Genetics of Populations, Volume 1: Genetic and Biometric Foundations. The University of Chicago Press. ISBN 0-226-91038-5.
- ↑ Terrall, M (2002). The Man who flattened the Earth: Maupertuis and the sciences in the Enlightenment. The University of Chicago Press. ISBN 978-0226793610.
- ↑ Wallace, A (1858). "On the Tendency of Species to form Varieties, and on the Perpetuation of Varieties and Species by Natural Means of Selection". Journal of the Proceedings of the Linnean Society of London. Zoology 3: 53–62. doi:10.1098/rsnr.2006.0171. http://darwin-online.org.uk/content/frameset?itemID=F350&viewtype=text&pageseq=1. Retrieved 2007-05-13.
- ↑ Darwin, Charles (1872). "Effects of the increased Use and Disuse of Parts, as controlled by Natural Selection". The Origin of Species. 6th edition, p. 108. John Murray. http://darwin-online.org.uk/content/frameset?viewtype=text&itemID=F391&pageseq=136. Retrieved 2007-12-28.
- ↑ Leakey, Richard E.; Darwin, Charles (1979). The illustrated origin of species. London: Faber. ISBN 0-571-14586-8. p. 17-18
- ↑ Ghiselin, Michael T. (September/October 1994). "Nonsense in schoolbooks: 'The Imaginary Lamarck'". The Textbook Letter. The Textbook League. http://www.textbookleague.org/54marck.htm. Retrieved 2008-01-23.
- ↑ Magner, Lois N. (2002). A History of the Life Sciences (Third ed.). Marcel Dekker, CRC Press. ISBN 9780203911006. http://books.google.com/?id=YKJ6gVYbrGwC&printsec=frontcover#v=onepage&q.
- ↑ Weiling F (1991). "Historical study: Johann Gregor Mendel 1822–1884". Am. J. Med. Genet. 40 (1): 1–25; discussion 26. doi:10.1002/ajmg.1320400103. PMID 1887835.
- ↑ Quammen, D. (2006). The reluctant Mr. Darwin: An intimate portrait of Charles Darwin and the making of his theory of evolution. New York, NY: W.W. Norton & Company.
- ↑ Bowler, Peter J. (1989). The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society. Baltimore: Johns Hopkins University Press. ISBN 978-0801838880.
- ↑ Pigliucci M (December 2007). "Do we need an extended evolutionary synthesis?". Evolution 61 (12): 2743–9. doi:10.1111/j.1558-5646.2007.00246.x. PMID 17924956.
- ↑ Winther RG (August 2008). "Systemic darwinism". Proc. Natl. Acad. Sci. U.S.A. 105 (33): 11833–8. doi:10.1073/pnas.0711445105. PMID 18697926.
- ↑ Sturm RA, Frudakis TN (2004). "Eye colour: portals into pigmentation genes and ancestry". Trends Genet. 20 (8): 327–32. doi:10.1016/j.tig.2004.06.010. PMID 15262401.
- ↑ 37.0 37.1 Pearson H (2006). "Genetics: what is a gene?". Nature 441 (7092): 398–401. doi:10.1038/441398a. PMID 16724031.
- ↑ Visscher PM, Hill WG, Wray NR (April 2008). "Heritability in the genomics era—concepts and misconceptions". Nat. Rev. Genet. 9 (4): 255–66. doi:10.1038/nrg2322. PMID 18319743.
- ↑ Oetting WS, Brilliant MH, King RA (1996). "The clinical spectrum of albinism in humans". Molecular medicine today 2 (8): 330–5. doi:10.1016/1357-4310(96)81798-9. PMID 8796918.
- ↑ Phillips PC (November 2008). "Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems". Nat. Rev. Genet. 9 (11): 855–67. doi:10.1038/nrg2452. PMID 18852697.
- ↑ 41.0 41.1 Wu R, Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits". Nat. Rev. Genet. 7 (3): 229–37. doi:10.1038/nrg1804. PMID 16485021.
- ↑ Richards EJ (May 2006). "Inherited epigenetic variation—revisiting soft inheritance". Nat. Rev. Genet. 7 (5): 395–401. doi:10.1038/nrg1834. PMID 16534512.
- ↑ 43.0 43.1 Harwood AJ; Harwood, J (1998). "Factors affecting levels of genetic diversity in natural populations". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353 (1366): 177–86. doi:10.1098/rstb.1998.0200. PMID 9533122.
- ↑ Draghi J, Turner P (2006). "DNA secretion and gene-level selection in bacteria". Microbiology (Reading, Engl.) 152 (Pt 9): 2683–8. doi:10.1099/mic.0.29013-0. PMID 16946263.
*Mallet J (2007). "Hybrid speciation". Nature 446 (7133): 279–83. doi:10.1038/nature05706. PMID 17361174. - ↑ Butlin RK, Tregenza T (1998). "Levels of genetic polymorphism: marker loci versus quantitative traits". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353 (1366): 187–98. doi:10.1098/rstb.1998.0201. PMID 9533123.
- ↑ Wetterbom A, Sevov M, Cavelier L, Bergström TF (2006). "Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution". J. Mol. Evol. 63 (5): 682–90. doi:10.1007/s00239-006-0045-7. PMID 17075697.
- ↑ 47.0 47.1 47.2 Bertram J (2000). "The molecular biology of cancer". Mol. Aspects Med. 21 (6): 167–223. doi:10.1016/S0098-2997(00)00007-8. PMID 11173079.
- ↑ 48.0 48.1 Aminetzach YT, Macpherson JM, Petrov DA (2005). "Pesticide resistance via transposition-mediated adaptive gene truncation in Drosophila". Science 309 (5735): 764–7. doi:10.1126/science.1112699. PMID 16051794.
- ↑ Burrus V, Waldor M (2004). "Shaping bacterial genomes with integrative and conjugative elements". Res. Microbiol. 155 (5): 376–86. doi:10.1016/j.resmic.2004.01.012. PMID 15207870.
- ↑ Sawyer SA, Parsch J, Zhang Z, Hartl DL (2007). "Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila". Proc. Natl. Acad. Sci. U.S.A. 104 (16): 6504–10. doi:10.1073/pnas.0701572104. PMID 17409186.
- ↑ Sniegowski P, Gerrish P, Johnson T, Shaver A (2000). "The evolution of mutation rates: separating causes from consequences". Bioessays 22 (12): 1057–66. doi:10.1002/1521-1878(200012)22:12<1057::AID-BIES3>3.0.CO;2-W. PMID 11084621.
- ↑ Drake JW, Holland JJ (1999). "Mutation rates among RNA viruses". Proc. Natl. Acad. Sci. U.S.A. 96 (24): 13910–3. doi:10.1073/pnas.96.24.13910. PMID 10570172. PMC 24164. http://www.pnas.org/content/96/24/13910.long.
- ↑ Holland J, Spindler K, Horodyski F, Grabau E, Nichol S, VandePol S (1982). "Rapid evolution of RNA genomes". Science 215 (4540): 1577–85. doi:10.1126/science.7041255. PMID 7041255.
- ↑ Hastings, P J; Lupski, JR; Rosenberg, SM; Ira, G (2009). "Mechanisms of change in gene copy number". Nature Reviews. Genetics 10 (8): 551–564. doi:10.1038/nrg2593. PMID 19597530.
- ↑ Carroll SB, Grenier J, Weatherbee SD (2005). From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Second Edition. Oxford: Blackwell Publishing. ISBN 1-4051-1950-0.
- ↑ Harrison P, Gerstein M (2002). "Studying genomes through the aeons: protein families, pseudogenes and proteome evolution". J Mol Biol 318 (5): 1155–74. doi:10.1016/S0022-2836(02)00109-2. PMID 12083509.
- ↑ Bowmaker JK (1998). "Evolution of colour vision in vertebrates". Eye (London, England) 12 (Pt 3b): 541–7. PMID 9775215.
- ↑ Gregory TR, Hebert PD (1999). "The modulation of DNA content: proximate causes and ultimate consequences". Genome Res. 9 (4): 317–24. doi:10.1101/gr.9.4.317 (inactive 2009-11-14). PMID 10207154. http://genome.cshlp.org/content/9/4/317.full.
- ↑ Hurles M (July 2004). "Gene duplication: the genomic trade in spare parts". PLoS Biol. 2 (7): E206. doi:10.1371/journal.pbio.0020206. PMID 15252449.
- ↑ Liu N, Okamura K, Tyler DM (2008). "The evolution and functional diversification of animal microRNA genes". Cell Res. 18 (10): 985–96. doi:10.1038/cr.2008.278. PMID 18711447. PMC 2712117. http://www.nature.com/cr/journal/v18/n10/full/cr2008278a.html.
- ↑ Siepel A (October 2009). "Darwinian alchemy: Human genes from noncoding DNA". Genome Res. 19 (10): 1693–5. doi:10.1101/gr.098376.109. PMID 19797681. PMC 2765273. http://genome.cshlp.org/content/19/10/1693.full.
- ↑ Orengo CA, Thornton JM (2005). "Protein families and their evolution-a structural perspective". Annu. Rev. Biochem. 74: 867–900. doi:10.1146/annurev.biochem.74.082803.133029. PMID 15954844.
- ↑ Long M, Betrán E, Thornton K, Wang W (November 2003). "The origin of new genes: glimpses from the young and old". Nat. Rev. Genet. 4 (11): 865–75. doi:10.1038/nrg1204. PMID 14634634.
- ↑ Wang M, Caetano-Anollés G (2009). "The evolutionary mechanics of domain organization in proteomes and the rise of modularity in the protein world". Structure 17 (1): 66–78. doi:10.1016/j.str.2008.11.008. PMID 19141283.
- ↑ Weissman KJ, Müller R (April 2008). "Protein-protein interactions in multienzyme megasynthetases". Chembiochem 9 (6): 826–48. doi:10.1002/cbic.200700751. PMID 18357594.
- ↑ Zhang J, Wang X, Podlaha O (2004). "Testing the chromosomal speciation hypothesis for humans and chimpanzees". Genome Res. 14 (5): 845–51. doi:10.1101/gr.1891104. PMID 15123584.
- ↑ Ayala FJ, Coluzzi M (2005). "Chromosome speciation: humans, Drosophila, and mosquitoes". Proc. Natl. Acad. Sci. U.S.A. 102 (Suppl 1): 6535–42. doi:10.1073/pnas.0501847102. PMID 15851677. PMC 1131864. http://www.pnas.org/content/102/suppl.1/6535.full.
- ↑ Hurst GD, Werren JH (2001). "The role of selfish genetic elements in eukaryotic evolution". Nat. Rev. Genet. 2 (8): 597–606. doi:10.1038/35084545. PMID 11483984.
- ↑ Häsler J, Strub K (2006). "Alu elements as regulators of gene expression". Nucleic Acids Res. 34 (19): 5491–7. doi:10.1093/nar/gkl706. PMID 17020921.
- ↑ Radding C (1982). "Homologous pairing and strand exchange in genetic recombination". Annu. Rev. Genet. 16: 405–37. doi:10.1146/annurev.ge.16.120182.002201. PMID 6297377.
- ↑ Agrawal AF (2006). "Evolution of sex: why do organisms shuffle their genotypes?". Curr. Biol. 16 (17): R696. doi:10.1016/j.cub.2006.07.063. PMID 16950096.
- ↑ Peters AD, Otto SP (2003). "Liberating genetic variance through sex". Bioessays 25 (6): 533–7. doi:10.1002/bies.10291. PMID 12766942.
- ↑ Goddard MR, Godfray HC, Burt A (2005). "Sex increases the efficacy of natural selection in experimental yeast populations". Nature 434 (7033): 636–40. doi:10.1038/nature03405. PMID 15800622.
- ↑ Fontaneto D, Herniou EA, Boschetti C (April 2007). "Independently evolving species in asexual bdelloid rotifers". PLoS Biol. 5 (4): e87. doi:10.1371/journal.pbio.0050087. PMID 17373857. Lay summary.
- ↑ Pouchkina-Stantcheva NN, McGee BM, Boschetti C (October 2007). "Functional divergence of former alleles in an ancient asexual invertebrate". Science 318 (5848): 268–71. doi:10.1126/science.1144363. PMID 17932297. Lay summary.
- ↑ Lien S, Szyda J, Schechinger B, Rappold G, Arnheim N (February 2000). "Evidence for heterogeneity in recombination in the human pseudoautosomal region: high resolution analysis by sperm typing and radiation-hybrid mapping". Am. J. Hum. Genet. 66 (2): 557–66. doi:10.1086/302754. PMID 10677316.
- ↑ Barton, N H (2000). "Genetic hitchhiking". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 355 (1403): 1553–1562. doi:10.1098/rstb.2000.0716. PMID 11127900. PMC 1692896. http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=1692896&blobtype=pdf. Retrieved 2009-09-23.
- ↑ Muller H (1964). "The relation of recombination to mutational advance". Mutat. Res. 106: 2–9. PMID 14195748.
- ↑ Charlesworth B, Charlesworth D (November 2000). "The degeneration of Y chromosomes". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 355 (1403): 1563–72. doi:10.1098/rstb.2000.0717. PMID 11127901.
- ↑ 80.0 80.1 Otto S (1 July 2003). "The advantages of segregation and the evolution of sex". Genetics 164 (3): 1099–118. PMID 12871918.
- ↑ Doncaster CP, Pound GE, Cox SJ (March 2000). "The ecological cost of sex". Nature 404 (6775): 281–5. doi:10.1038/35005078. PMID 10749210.
- ↑ Butlin R (April 2002). "Evolution of sex: The costs and benefits of sex: new insights from old asexual lineages". Nat. Rev. Genet. 3 (4): 311–7. doi:10.1038/nrg749. PMID 11967555.
- ↑ Salathé M, Kouyos RD, Bonhoeffer S (August 2008). "The state of affairs in the kingdom of the Red Queen". Trends Ecol. Evol. (Amst.) 23 (8): 439–45. doi:10.1016/j.tree.2008.04.010. PMID 18597889.
- ↑ Stoltzfus A (2006). "Mutationism and the dual causation of evolutionary change". Evol. Dev. 8 (3): 304–17. doi:10.1111/j.1525-142X.2006.00101.x. PMID 16686641.
- ↑ O'Neil, Dennis (2008). "Hardy-Weinberg Equilibrium Model". The synthetic theory of evolution: An introduction to modern evolutionary concepts and theories. Behavioral Sciences Department, Palomar College. http://anthro.palomar.edu/synthetic/synth_2.htm. Retrieved 2008-01-06.
- ↑ Bright, Kerry (2006). "Causes of evolution". Teach Evolution and Make It Relevant. National Science Foundation. http://evoled.dbs.umt.edu/lessons/causes.htm#hardy. Retrieved 2007-12-30.
- ↑ Packer C, Gilbert DA, Pusey AE, O'Brieni SJ. (June 1991). "A molecular genetic analysis of kinship and cooperation in African lions". Nature 351: 562–65. doi:10.1038/351562a0.
- ↑ Morjan C, Rieseberg L (2004). "How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles". Mol. Ecol. 13 (6): 1341–56. doi:10.1111/j.1365-294X.2004.02164.x. PMID 15140081.
- ↑ Su H, Qu L, He K, Zhang Z, Wang J, Chen Z, Gu H (2003). "The Great Wall of China: a physical barrier to gene flow?". Heredity 90 (3): 212–9. doi:10.1038/sj.hdy.6800237. PMID 12634804.
- ↑ Short RV (1975). "The contribution of the mule to scientific thought". J. Reprod. Fertil. Suppl. (23): 359–64. PMID 1107543.
- ↑ Gross B, Rieseberg L (2005). "The ecological genetics of homoploid hybrid speciation". J. Hered. 96 (3): 241–52. doi:10.1093/jhered/esi026. PMID 15618301.
- ↑ Burke JM, Arnold ML (2001). "Genetics and the fitness of hybrids". Annu. Rev. Genet. 35: 31–52. doi:10.1146/annurev.genet.35.102401.085719. PMID 11700276.
- ↑ Vrijenhoek RC (2006). "Polyploid hybrids: multiple origins of a treefrog species". Curr. Biol. 16 (7): R245. doi:10.1016/j.cub.2006.03.005. PMID 16581499.
- ↑ Wendel J (2000). "Genome evolution in polyploids". Plant Mol. Biol. 42 (1): 225–49. doi:10.1023/A:1006392424384. PMID 10688139.
- ↑ 95.0 95.1 Sémon M, Wolfe KH (2007). "Consequences of genome duplication". Curr Opin Genet Dev 17 (6): 505–12. doi:10.1016/j.gde.2007.09.007. PMID 18006297.
- ↑ Comai L (2005). "The advantages and disadvantages of being polyploid". Nat. Rev. Genet. 6 (11): 836–46. doi:10.1038/nrg1711. PMID 16304599.
- ↑ Soltis P, Soltis D (June 2000). "The role of genetic and genomic attributes in the success of polyploids". Proc. Natl. Acad. Sci. U.S.A. 97 (13): 7051–7. doi:10.1073/pnas.97.13.7051. PMID 10860970.
- ↑ Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF (2003). "Lateral gene transfer and the origins of prokaryotic groups". Annu Rev Genet 37: 283–328. doi:10.1146/annurev.genet.37.050503.084247. PMID 14616063.
- ↑ Kondo N, Nikoh N, Ijichi N, Shimada M, Fukatsu T (2002). "Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect". Proc. Natl. Acad. Sci. U.S.A. 99 (22): 14280–5. doi:10.1073/pnas.222228199. PMID 12386340.
- ↑ Sprague G (1991). "Genetic exchange between kingdoms". Curr. Opin. Genet. Dev. 1 (4): 530–3. doi:10.1016/S0959-437X(05)80203-5. PMID 1822285.
- ↑ Gladyshev EA, Meselson M, Arkhipova IR (May 2008). "Massive horizontal gene transfer in bdelloid rotifers". Science 320 (5880): 1210–3. doi:10.1126/science.1156407. PMID 18511688.
- ↑ Baldo A, McClure M (1 September 1999). "Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts". J. Virol. 73 (9): 7710–21. PMID 10438861.
- ↑ 103.0 103.1 Poole A, Penny D (2007). "Evaluating hypotheses for the origin of eukaryotes". Bioessays 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354.
- ↑ Whitlock M (1 June 2003). "Fixation probability and time in subdivided populations". Genetics 164 (2): 767–79. PMID 12807795.
- ↑ Ohta T (2002). "Near-neutrality in evolution of genes and gene regulation". Proc. Natl. Acad. Sci. U.S.A. 99 (25): 16134–7. doi:10.1073/pnas.252626899. PMID 12461171. PMC 138577. http://www.pnas.org/cgi/content/abstract/252626899v1.
- ↑ 106.0 106.1 106.2 Hurst LD (February 2009). "Fundamental concepts in genetics: genetics and the understanding of selection". Nat. Rev. Genet. 10 (2): 83–93. doi:10.1038/nrg2506. PMID 19119264.
- ↑ 107.0 107.1 107.2 Orr HA (August 2009). "Fitness and its role in evolutionary genetics". Nat. Rev. Genet. 10 (8): 531–9. doi:10.1038/nrg2603. PMID 19546856.
- ↑ Haldane J (1959). "The theory of natural selection today". Nature 183 (4663): 710–3. doi:10.1038/183710a0. PMID 13644170.
- ↑ Lande R, Arnold SJ (1983). "The measurement of selection on correlated characters". Evolution 37 (6): 1210–26. doi:10.2307/2408842. http://jstor.org/stable/2408842.
- ↑ Goldberg, Emma E; Igić, B (2008). "On phylogenetic tests of irreversible evolution". Evolution 62 (11): 2727–2741. doi:10.1111/j.1558-5646.2008.00505.x. PMID 18764918.
- ↑ Collin, Rachel; Miglietta, MP (2008). "Reversing opinions on Dollo's Law". Trends in Ecology & Evolution 23 (11): 602–609. doi:10.1016/j.tree.2008.06.013. PMID 18814933.
- ↑ Hoekstra H, Hoekstra J, Berrigan D, Vignieri S, Hoang A, Hill C, Beerli P, Kingsolver J (July 2001). "Strength and tempo of directional selection in the wild". Proc. Natl. Acad. Sci. U.S.A. 98 (16): 9157–60. doi:10.1073/pnas.161281098. PMID 11470913.
- ↑ Felsenstein (1 November 1979). "Excursions along the Interface between Disruptive and Stabilizing Selection". Genetics 93 (3): 773–95. PMID 17248980.
- ↑ Andersson M, Simmons L (2006). "Sexual selection and mate choice". Trends Ecol. Evol. (Amst.) 21 (6): 296–302. doi:10.1016/j.tree.2006.03.015. PMID 16769428.
- ↑ Kokko H, Brooks R, McNamara J, Houston A (2002). "The sexual selection continuum". Proc. Biol. Sci. 269 (1498): 1331–40. doi:10.1098/rspb.2002.2020. PMID 12079655.
- ↑ Hunt J, Brooks R, Jennions M, Smith M, Bentsen C, Bussière L (2004). "High-quality male field crickets invest heavily in sexual display but die young". Nature 432 (7020): 1024–7. doi:10.1038/nature03084. PMID 15616562.
- ↑ Odum, EP (1971) Fundamentals of ecology, third edition, Saunders New York
- ↑ 118.0 118.1 Gould SJ (February 1998). "Gulliver's further travels: the necessity and difficulty of a hierarchical theory of selection". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353 (1366): 307–14. doi:10.1098/rstb.1998.0211. PMID 9533127.
- ↑ Mayr E (1997). "The objects of selection". Proc. Natl. Acad. Sci. U.S.A. 94 (6): 2091–4. doi:10.1073/pnas.94.6.2091. PMID 9122151.
- ↑ Maynard Smith J (1998). "The units of selection". Novartis Found. Symp. 213: 203–11; discussion 211–7. PMID 9653725.
- ↑ Hickey DA (1992). "Evolutionary dynamics of transposable elements in prokaryotes and eukaryotes". Genetica 86 (1–3): 269–74. doi:10.1007/BF00133725. PMID 1334911.
- ↑ Gould SJ, Lloyd EA (1999). "Individuality and adaptation across levels of selection: how shall we name and generalize the unit of Darwinism?". Proc. Natl. Acad. Sci. U.S.A. 96 (21): 11904–9. doi:10.1073/pnas.96.21.11904. PMID 10518549.
- ↑ Lande R (1989). "Fisherian and Wrightian theories of speciation". Genome 31 (1): 221–7. PMID 2687093.
- ↑ Otto S, Whitlock M (1 June 1997). "The probability of fixation in populations of changing size". Genetics 146 (2): 723–33. PMID 9178020.
- ↑ Charlesworth B (March 2009). "Fundamental concepts in genetics: Effective population size and patterns of molecular evolution and variation". Nat. Rev. Genet. 10 (3): 195–205. doi:10.1038/nrg2526. PMID 19204717.
- ↑ Protas, Meredith; Conrad, M; Gross, JB; Tabin, C; Borowsky, R (2007). "Regressive evolution in the Mexican cave tetra, Astyanax mexicanus". Current Biology 17 (5): 452–454. doi:10.1016/j.cub.2007.01.051. PMID 17306543. PMC 2570642. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=17306543#R2.
- ↑ Mitchell-Olds, Thomas; Willis, JH; Goldstein, DB (2007). "Which evolutionary processes influence natural genetic variation for phenotypic traits?". Nature Reviews Genetics 8 (11): 845–856. doi:10.1038/nrg2207. PMID 17943192.
- ↑ Nei M (2005). "Selectionism and neutralism in molecular evolution". Mol. Biol. Evol. 22 (12): 2318–42. doi:10.1093/molbev/msi242. PMID 16120807.
- ↑ Kimura M (1991). "The neutral theory of molecular evolution: a review of recent evidence". Jpn. J. Genet. 66 (4): 367–86. doi:10.1266/jjg.66.367. PMID 1954033. http://www.jstage.jst.go.jp/article/jjg/66/4/66_367/_article.
- ↑ Kimura M (1989). "The neutral theory of molecular evolution and the world view of the neutralists". Genome 31 (1): 24–31. PMID 2687096.
- ↑ Kreitman M (August 1996). "The neutral theory is dead. Long live the neutral theory". Bioessays 18 (8): 678–83; discussion 683. doi:10.1002/bies.950180812. PMID 8760341. http://www.cs.ucsb.edu/~ambuj/Courses/bioinformatics/neutral-theory.
- ↑ Leigh E.G. (Jr) (2007). "Neutral theory: a historical perspective.". Journal of Evolutionary Biology 20 (6): 2075–91. doi:10.1111/j.1420-9101.2007.01410.x. PMID 17956380.
- ↑ 133.0 133.1 133.2 Scott EC, Matzke NJ (May 2007). "Biological design in science classrooms". Proc. Natl. Acad. Sci. U.S.A. 104 Suppl 1: 8669–76. doi:10.1073/pnas.0701505104. PMID 17494747.
- ↑ Hendry AP, Kinnison MT (2001). "An introduction to microevolution: rate, pattern, process". Genetica 112–113: 1–8. doi:10.1023/A:1013368628607. PMID 11838760.
- ↑ Leroi AM (2000). "The scale independence of evolution". Evol. Dev. 2 (2): 67–77. doi:10.1046/j.1525-142x.2000.00044.x. PMID 11258392.
- ↑ Gould 2002, pp. 657–8
- ↑ Gould SJ (July 1994). "Tempo and mode in the macroevolutionary reconstruction of Darwinism". Proc. Natl. Acad. Sci. U.S.A. 91 (15): 6764–71. doi:10.1073/pnas.91.15.6764. PMID 8041695. PMC 44281. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=8041695.
- ↑ Jablonski, D. (2000). "Micro- and macroevolution: scale and hierarchy in evolutionary biology and paleobiology". Paleobiology 26 (sp4): 15–52. doi:10.1666/0094-8373(2000)26[15:MAMSAH]2.0.CO;2. http://www.bioone.org/perlserv/?request=get-abstract.
- ↑ Michael J. Dougherty. Is the human race evolving or devolving? Scientific American July 20, 1998.
- ↑ TalkOrigins Archive response to Creationist claims – Claim CB932: Evolution of degenerate forms
- ↑ Carroll SB (2001). "Chance and necessity: the evolution of morphological complexity and diversity". Nature 409 (6823): 1102–9. doi:10.1038/35059227. PMID 11234024.
- ↑ Whitman W, Coleman D, Wiebe W (1998). "Prokaryotes: the unseen majority". Proc Natl Acad Sci U S a 95 (12): 6578–83. doi:10.1073/pnas.95.12.6578. PMID 9618454.
- ↑ 143.0 143.1 Schloss P, Handelsman J (December 2004). "Status of the microbial census". Microbiol Mol Biol Rev 68 (4): 686–91. doi:10.1128/MMBR.68.4.686-691.2004. PMID 15590780.
- ↑ Nealson K (1999). "Post-Viking microbiology: new approaches, new data, new insights". Orig Life Evol Biosph 29 (1): 73–93. doi:10.1023/A:1006515817767. PMID 11536899.
- ↑ Buckling A, Craig Maclean R, Brockhurst MA, Colegrave N (February 2009). "The Beagle in a bottle". Nature 457 (7231): 824–9. doi:10.1038/nature07892. PMID 19212400.
- ↑ Elena SF, Lenski RE (June 2003). "Evolution experiments with microorganisms: the dynamics and genetic bases of adaptation". Nat. Rev. Genet. 4 (6): 457–69. doi:10.1038/nrg1088. PMID 12776215.
- ↑ Williams, George C. 1966. Adaptation and natural selection: a critique of some current evolutionary thought. Princeton. "Evolutionary adaptation is a phenomenon of pervasive importance in biology." p5
- ↑ Mayr, Ernst 1982. The growth of biological thought. Harvard. p483: "Adaptation... could no longer be considered a static condition, a product of a creative past, and became instead a continuing dynamic process."
- ↑ The Oxford Dictionary of Science defines adaptation as "Any change in the structure or functioning of an organism that makes it better suited to its environment".
- ↑ Orr H (2005). "The genetic theory of adaptation: a brief history". Nat. Rev. Genet. 6 (2): 119–27. doi:10.1038/nrg1523. PMID 15716908.
- ↑ Dobzhansky T. 1968. On some fundamental concepts of evolutionary biology. Evolutionary biology 2, 1–34.
- ↑ Dobzhansky T. 1970. Genetics of the evolutionary process. Columbia, N.Y. p4–6, 79–82, 84–87
- ↑ Dobzhansky T. 1956. Genetics of natural populations XXV. Genetic changes in populations of Drosophila pseudoobscura and Drosphila persimilis in some locations in California. Evolution 10, 82–92.
- ↑ Nakajima A, Sugimoto Y, Yoneyama H, Nakae T (2002). "High-level fluoroquinolone resistance in Pseudomonas aeruginosa due to interplay of the MexAB-OprM efflux pump and the DNA gyrase mutation". Microbiol. Immunol. 46 (6): 391–5. PMID 12153116. http://www.jstage.jst.go.jp/article/mandi/46/6/46_391/_article/-char/en.
- ↑ Blount ZD, Borland CZ, Lenski RE (June 2008). "Inaugural Article: Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli". Proc. Natl. Acad. Sci. U.S.A. 105 (23): 7899–906. doi:10.1073/pnas.0803151105. PMID 18524956. PMC 2430337. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=18524956.
- ↑ Okada H, Negoro S, Kimura H, Nakamura S (1983). "Evolutionary adaptation of plasmid-encoded enzymes for degrading nylon oligomers". Nature 306 (5939): 203–6. doi:10.1038/306203a0. PMID 6646204.
- ↑ Ohno S (April 1984). "Birth of a unique enzyme from an alternative reading frame of the preexisted, internally repetitious coding sequence". Proc. Natl. Acad. Sci. U.S.A. 81 (8): 2421–5. doi:10.1073/pnas.81.8.2421. PMID 6585807. PMC 345072. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=6585807.
- ↑ Copley SD (June 2000). "Evolution of a metabolic pathway for degradation of a toxic xenobiotic: the patchwork approach". Trends Biochem. Sci. 25 (6): 261–5. doi:10.1016/S0968-0004(00)01562-0. PMID 10838562.
- ↑ Crawford RL, Jung CM, Strap JL (October 2007). "The recent evolution of pentachlorophenol (PCP)-4-monooxygenase (PcpB) and associated pathways for bacterial degradation of PCP". Biodegradation 18 (5): 525–39. doi:10.1007/s10532-006-9090-6. PMID 17123025.
- ↑ Colegrave N, Collins S (May 2008). "Experimental evolution: experimental evolution and evolvability". Heredity 100 (5): 464–70. doi:10.1038/sj.hdy.6801095. PMID 18212804. http://www.nature.com/hdy/journal/v100/n5/full/6801095a.html.
- ↑ Kirschner M, Gerhart J (July 1998). "Evolvability". Proc. Natl. Acad. Sci. U.S.A. 95 (15): 8420–7. doi:10.1073/pnas.95.15.8420. PMID 9671692.
- ↑ 162.0 162.1 Bejder L, Hall BK (2002). "Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss". Evol. Dev. 4 (6): 445–58. doi:10.1046/j.1525-142X.2002.02033.x. PMID 12492145.
- ↑ Young, Nathan M.; Hallgrímsson, B (2005). "Serial homology and the evolution of mammalian limb covariation structure". Evolution 59 (12): 2691. doi:10.1554/05-233.1. PMID 16526515. http://www.bioone.org/doi/abs/10.1554/05-233.1. Retrieved 2009-09-24.
- ↑ 164.0 164.1 Penny D, Poole A (1999). "The nature of the last universal common ancestor". Curr. Opin. Genet. Dev. 9 (6): 672–77. doi:10.1016/S0959-437X(99)00020-9. PMID 10607605.
- ↑ Hall, Brian K (2003). "Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution". Biological Reviews of the Cambridge Philosophical Society 78 (3): 409–433. doi:10.1017/S1464793102006097. PMID 14558591.
- ↑ Shubin, Neil; Tabin, C; Carroll, S (2009). "Deep homology and the origins of evolutionary novelty". Nature 457 (7231): 818–823. doi:10.1038/nature07891. PMID 19212399.
- ↑ 167.0 167.1 167.2 Fong D, Kane T, Culver D (1995). "Vestigialization and Loss of Nonfunctional Characters". Ann. Rev. Ecol. Syst. 26: 249–68. doi:10.1146/annurev.es.26.110195.001341.
- ↑ Zhang Z, Gerstein M (August 2004). "Large-scale analysis of pseudogenes in the human genome". Curr. Opin. Genet. Dev. 14 (4): 328–35. doi:10.1016/j.gde.2004.06.003. PMID 15261647.
- ↑ Jeffery WR (2005). "Adaptive evolution of eye degeneration in the Mexican blind cavefish". J. Hered. 96 (3): 185–96. doi:10.1093/jhered/esi028. PMID 15653557.
- ↑ Maxwell EE, Larsson HC (2007). "Osteology and myology of the wing of the Emu (Dromaius novaehollandiae), and its bearing on the evolution of vestigial structures". J. Morphol. 268 (5): 423–41. doi:10.1002/jmor.10527. PMID 17390336.
- ↑ Silvestri AR, Singh I (2003). "The unresolved problem of the third molar: would people be better off without it?". Journal of the American Dental Association (1939) 134 (4): 450–5. doi:10.1146/annurev.es.26.110195.001341. PMID 12733778. http://jada.ada.org/cgi/content/full/134/4/450.
- ↑ Darwin, Charles. (1872) The Expression of the Emotions in Man and Animals John Murray, London.
- ↑ Peter Gray (2007). Psychology (fifth ed.). Worth Publishers. pp. 66. ISBN 0-7167-0617-2.
- ↑ Behavior Development in Infants (via Google Books) by Evelyn Dewey, citing a study "Reflexes and other motor activities in newborn infants: a report of 125 cases as a preliminary study of infant behavior" published in the Bull. Neurol. Inst. New York, 1932, Vol. 2, pp. 1-56.
- ↑ Jerry Coyne (2009). Why Evolution is True. Penguin Group. pp. 85–86. ISBN 978-0-670-02053.
- ↑ Anthony Stevens (1982). Archetype: A Natural History of the Self. Routledge & Kegan Paul. pp. 87. ISBN 0-7100-0980-1.
- ↑ 177.0 177.1 Gould 2002, pp. 1235–6
- ↑ Pallen, Mark J.; Matzke, NJ (2006-10). "From The Origin of Species to the origin of bacterial flagella". Nat Rev Micro 4 (10): 784–790. doi:10.1038/nrmicro1493. PMID 16953248. http://home.planet.nl/~gkorthof/pdf/Pallen_Matzke.pdf. Retrieved 2009-09-18.
- ↑ Clements, Abigail; Bursac, D; Gatsos, X; Perry, AJ; Civciristov, S; Celik, N; Likic, VA; Poggio, S et al. (2009). "The reducible complexity of a mitochondrial molecular machine". Proceedings of the National Academy of Sciences 106 (37): 15791–15795. doi:10.1073/pnas.0908264106. PMID 19717453. PMC 2747197. http://www.pnas.org/content/106/37/15791.long. Retrieved 2009-09-18.
- ↑ Piatigorsky J, Kantorow M, Gopal-Srivastava R, Tomarev SI (1994). "Recruitment of enzymes and stress proteins as lens crystallins". EXS 71: 241–50. PMID 8032155.
- ↑ Wistow G (August 1993). "Lens crystallins: gene recruitment and evolutionary dynamism". Trends Biochem. Sci. 18 (8): 301–6. doi:10.1016/0968-0004(93)90041-K. PMID 8236445.
- ↑ Hardin G (April 1960). "The competitive exclusion principle". Science 131: 1292–7. doi:10.1126/science.131.3409.1292. PMID 14399717. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=14399717.
- ↑ Kocher TD (April 2004). "Adaptive evolution and explosive speciation: the cichlid fish model". Nat. Rev. Genet. 5 (4): 288–98. doi:10.1038/nrg1316. PMID 15131652. http://hcgs.unh.edu/staff/kocher/pdfs/Kocher2004.pdf.
- ↑ Johnson NA, Porter AH (2001). "Toward a new synthesis: population genetics and evolutionary developmental biology". Genetica 112–113: 45–58. doi:10.1023/A:1013371201773. PMID 11838782.
- ↑ Baguñà J, Garcia-Fernàndez J (2003). "Evo-Devo: the long and winding road". Int. J. Dev. Biol. 47 (7–8): 705–13. PMID 14756346. http://www.ijdb.ehu.es/web/paper.php?doi=14756346.
*Love AC. (2003). "Evolutionary Morphology, Innovation, and the Synthesis of Evolutionary and Developmental Biology". Biology and Philosophy 18 (2): 309–345. doi:10.1023/A:1023940220348. http://philsci-archive.pitt.edu/archive/00000375/00/LondonPaper.doc. - ↑ Allin EF (1975). "Evolution of the mammalian middle ear". J. Morphol. 147 (4): 403–37. doi:10.1002/jmor.1051470404. PMID 1202224.
- ↑ Harris MP, Hasso SM, Ferguson MW, Fallon JF (2006). "The development of archosaurian first-generation teeth in a chicken mutant". Curr. Biol. 16 (4): 371–7. doi:10.1016/j.cub.2005.12.047. PMID 16488870.
- ↑ Carroll SB (July 2008). "Evo-devo and an expanding evolutionary synthesis: a genetic theory of morphological evolution". Cell 134 (1): 25–36. doi:10.1016/j.cell.2008.06.030. PMID 18614008.
- ↑ Wade MJ (2007). "The co-evolutionary genetics of ecological communities". Nat. Rev. Genet. 8 (3): 185–95. doi:10.1038/nrg2031. PMID 17279094.
- ↑ Geffeney S, Brodie ED, Ruben PC, Brodie ED (2002). "Mechanisms of adaptation in a predator-prey arms race: TTX-resistant sodium channels". Science 297 (5585): 1336–9. doi:10.1126/science.1074310. PMID 12193784.
*Brodie ED, Ridenhour BJ, Brodie ED (2002). "The evolutionary response of predators to dangerous prey: hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts". Evolution 56 (10): 2067–82. PMID 12449493.
*Sean B. Carroll (December 21, 2009). "Remarkable Creatures – Clues to Toxins in Deadly Delicacies of the Animal Kingdom". New York Times. http://www.nytimes.com/2009/12/22/science/22creature.html?hpw. - ↑ Sachs J (2006). "Cooperation within and among species". J. Evol. Biol. 19 (5): 1415–8; discussion 1426–36. doi:10.1111/j.1420-9101.2006.01152.x. PMID 16910971.
*Nowak M (2006). "Five rules for the evolution of cooperation". Science 314 (5805): 1560–3. doi:10.1126/science.1133755. PMID 17158317. - ↑ Paszkowski U (2006). "Mutualism and parasitism: the yin and yang of plant symbioses". Curr. Opin. Plant Biol. 9 (4): 364–70. doi:10.1016/j.pbi.2006.05.008. PMID 16713732.
- ↑ Hause B, Fester T (2005). "Molecular and cell biology of arbuscular mycorrhizal symbiosis". Planta 221 (2): 184–96. doi:10.1007/s00425-004-1436-x. PMID 15871030.
- ↑ Reeve HK, Hölldobler B (2007). "The emergence of a superorganism through intergroup competition". Proc Natl Acad Sci U S A. 104 (23): 9736–40. doi:10.1073/pnas.0703466104. PMID 17517608.
- ↑ Axelrod R, Hamilton W (2005). "The evolution of cooperation". Science 211 (4489): 1390–6. doi:10.1126/science.7466396. PMID 7466396.
- ↑ Wilson EO, Hölldobler B (2005). "Eusociality: origin and consequences". Proc. Natl. Acad. Sci. U.S.A. 102 (38): 13367–71. doi:10.1073/pnas.0505858102. PMID 16157878.
- ↑ 197.0 197.1 Gavrilets S (2003). "Perspective: models of speciation: what have we learned in 40 years?". Evolution 57 (10): 2197–215. doi:10.1554/02-727. PMID 14628909.
- ↑ De Queiroz K (December 2007). "Species concepts and species delimitation". Syst. Biol. 56 (6): 879–86. doi:10.1080/10635150701701083. PMID 18027281.
- ↑ Fraser C, Alm EJ, Polz MF, Spratt BG, Hanage WP (February 2009). "The bacterial species challenge: making sense of genetic and ecological diversity". Science 323 (5915): 741–6. doi:10.1126/science.1159388. PMID 19197054.
- ↑ 200.0 200.1 de Queiroz K (May 2005). "Ernst Mayr and the modern concept of species". Proc. Natl. Acad. Sci. U.S.A. 102 (Suppl 1): 6600–7. doi:10.1073/pnas.0502030102. PMID 15851674. PMC 1131873. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=15851674.
- ↑ Rice, W.R.; Hostert (1993). "Laboratory experiments on speciation: what have we learned in 40 years". Evolution 47 (6): 1637–1653. doi:10.2307/2410209.
*Jiggins CD, Bridle JR (2004). "Speciation in the apple maggot fly: a blend of vintages?". Trends Ecol. Evol. (Amst.) 19 (3): 111–4. doi:10.1016/j.tree.2003.12.008. PMID 16701238.
*Boxhorn, J (1995). "Observed Instances of Speciation". TalkOrigins Archive. http://www.talkorigins.org/faqs/faq-speciation.html. Retrieved 2008-12-26.
*Weinberg JR, Starczak VR, Jorg, D (1992). "Evidence for Rapid Speciation Following a Founder Event in the Laboratory". Evolution 46 (4): 1214–20. doi:10.2307/2409766. http://jstor.org/stable/2409766. - ↑ Herrel, A.; Huyghe, K.; Vanhooydonck, B.; Backeljau, T.; Breugelmans, K.; Grbac, I.; Van Damme, R.; Irschick, D.J. (2008). "Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource". Proceedings of the National Academy of Sciences 105 (12): 4792–5. doi:10.1073/pnas.0711998105. PMID 18344323.
- ↑ Losos, J.B. Warhelt, K.I. Schoener, T.W. (1997). "Adaptive differentiation following experimental island colonization in Anolis lizards". Nature 387 (6628): 70–3. doi:10.1038/387070a0.
- ↑ Hoskin CJ, Higgle M, McDonald KR, Moritz C (2005). "Reinforcement drives rapid allopatric speciation". Nature 437 (7063): 1353–356. doi:10.1038/nature04004. PMID 16251964.
- ↑ Templeton AR (1 April 1980). "The theory of speciation via the founder principle". Genetics 94 (4): 1011–38. PMID 6777243. PMC 1214177. http://www.genetics.org/cgi/reprint/94/4/1011.
- ↑ Antonovics J (2006). "Evolution in closely adjacent plant populations X: long-term persistence of prereproductive isolation at a mine boundary". Heredity 97 (1): 33–7. doi:10.1038/sj.hdy.6800835. PMID 16639420. http://www.nature.com/hdy/journal/v97/n1/full/6800835a.html.
- ↑ Nosil P, Crespi B, Gries R, Gries G (2007). "Natural selection and divergence in mate preference during speciation". Genetica 129 (3): 309–27. doi:10.1007/s10709-006-0013-6. PMID 16900317.
- ↑ Savolainen V, Anstett M-C, Lexer C, Hutton I, Clarkson JJ, Norup MV, Powell MP, Springate D, Salamin N, Baker WJr (2006). "Sympatric speciation in palms on an oceanic island". Nature 441 (7090): 210–3. doi:10.1038/nature04566. PMID 16467788.
*Barluenga M, Stölting KN, Salzburger W, Muschick M, Meyer A (2006). "Sympatric speciation in Nicaraguan crater lake cichlid fish". Nature 439 (7077): 719–23. doi:10.1038/nature04325. PMID 16467837. - ↑ Gavrilets S (2006). "The Maynard Smith model of sympatric speciation". J. Theor. Biol. 239 (2): 172–82. doi:10.1016/j.jtbi.2005.08.041. PMID 16242727.
- ↑ Wood TE, Takebayashi N, Barker MS, Mayrose I, Greenspoon PB, Rieseberg LH (August 2009). "The frequency of polyploid speciation in vascular plants". Proc. Natl. Acad. Sci. U.S.A. 106 (33): 13875–9. doi:10.1073/pnas.0811575106. PMID 19667210.
- ↑ Hegarty Mf, Hiscock SJ (2008). "Genomic clues to the evolutionary success of polyploid plants". Current Biology 18 (10): 435–44. doi:10.1016/j.cub.2008.03.043. PMID 18492478.
- ↑ Jakobsson M, Hagenblad J, Tavaré S (2006). "A unique recent origin of the allotetraploid species Arabidopsis suecica: Evidence from nuclear DNA markers". Mol. Biol. Evol. 23 (6): 1217–31. doi:10.1093/molbev/msk006. PMID 16549398.
- ↑ Säll T, Jakobsson M, Lind-Halldén C, Halldén C (2003). "Chloroplast DNA indicates a single origin of the allotetraploid Arabidopsis suecica". J. Evol. Biol. 16 (5): 1019–29. doi:10.1046/j.1420-9101.2003.00554.x. PMID 14635917.
- ↑ Bomblies K, Weigel D (2007). "Arabidopsis-a model genus for speciation". Curr Opin Genet Dev 17 (6): 500–4. doi:10.1016/j.gde.2007.09.006. PMID 18006296.
- ↑ Niles Eldredge and Stephen Jay Gould, 1972. "Punctuated equilibria: an alternative to phyletic gradualism" In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82–115. Reprinted in N. Eldredge Time frames. Princeton: Princeton Univ. Press. 1985
- ↑ Gould SJ (1994). "Tempo and mode in the macroevolutionary reconstruction of Darwinism". Proc. Natl. Acad. Sci. U.S.A. 91 (15): 6764–71. doi:10.1073/pnas.91.15.6764. PMID 8041695.
- ↑ Benton MJ (1995). "Diversification and extinction in the history of life". Science 268 (5207): 52–8. doi:10.1126/science.7701342. PMID 7701342.
- ↑ Raup DM (1986). "Biological extinction in Earth history". Science 231: 1528–33. doi:10.1126/science.11542058. PMID 11542058.
- ↑ Avise JC, Hubbell SP, Ayala FJ. (August 2008). "In the light of evolution II: Biodiversity and extinction". Proc. Natl. Acad. Sci. U.S.A. 105 (Suppl 1): 11453–7. doi:10.1073/pnas.0802504105. PMID 18695213. PMC 2556414. http://www.pnas.org/content/105/suppl.1/11453.full.
- ↑ 220.0 220.1 220.2 Raup DM (1994). "The role of extinction in evolution". Proc. Natl. Acad. Sci. U.S.A. 91 (15): 6758–63. doi:10.1073/pnas.91.15.6758. PMID 8041694.
- ↑ Novacek MJ, Cleland EE (2001). "The current biodiversity extinction event: scenarios for mitigation and recovery". Proc. Natl. Acad. Sci. U.S.A. 98 (10): 5466–70. doi:10.1073/pnas.091093698. PMID 11344295.
- ↑ Pimm S, Raven P, Peterson A, Sekercioglu CH, Ehrlich PR (2006). "Human impacts on the rates of recent, present, and future bird extinctions". Proc. Natl. Acad. Sci. U.S.A. 103 (29): 10941–6. doi:10.1073/pnas.0604181103. PMID 16829570.
*Barnosky AD, Koch PL, Feranec RS, Wing SL, Shabel AB (2004). "Assessing the causes of late Pleistocene extinctions on the continents". Science 306 (5693): 70–5. doi:10.1126/science.1101476. PMID 15459379. - ↑ Lewis OT (2006). "Climate change, species-area curves and the extinction crisis". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 361 (1465): 163–71. doi:10.1098/rstb.2005.1712. PMID 16553315.
- ↑ Jablonski D (May 2001). "Lessons from the past: evolutionary impacts of mass extinctions". Proc. Natl. Acad. Sci. U.S.A. 98 (10): 5393–8. doi:10.1073/pnas.101092598. PMID 11344284.
- ↑ Isaak, Mark (2005). "Claim CB090: Evolution without abiogenesis". TalkOrigins Archive. http://www.talkorigins.org/indexcc/CB/CB090.html. Retrieved 2008-12-26.
- ↑ Peretó J (2005). "Controversies on the origin of life" (PDF). Int. Microbiol. 8 (1): 23–31. PMID 15906258. http://www.im.microbios.org/0801/0801023.pdf.
- ↑ Luisi PL, Ferri F, Stano P (2006). "Approaches to semi-synthetic minimal cells: a review". Naturwissenschaften 93 (1): 1–13. doi:10.1007/s00114-005-0056-z. PMID 16292523.
- ↑ Trevors JT, Abel DL (2004). "Chance and necessity do not explain the origin of life". Cell Biol. Int. 28 (11): 729–39. doi:10.1016/j.cellbi.2004.06.006. PMID 15563395.Forterre P, Benachenhou-Lahfa N, Confalonieri F, Duguet M, Elie C, Labedan B (1992). "The nature of the last universal ancestor and the root of the tree of life, still open questions". BioSystems 28 (1–3): 15–32. doi:10.1016/0303-2647(92)90004-I. PMID 1337989.
- ↑ Joyce GF (2002). "The antiquity of RNA-based evolution". Nature 418 (6894): 214–21. doi:10.1038/418214a. PMID 12110897.
- ↑ Trevors JT, Psenner R (2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". FEMS Microbiol. Rev. 25 (5): 573–82. doi:10.1111/j.1574-6976.2001.tb00592.x. PMID 11742692.
- ↑ Theobald, DL. (2010). "A formal test of the theory of universal common ancestry". Nature 465 (7295): 219–22. doi:10.1038/nature09014. PMID 20463738.
- ↑ Bapteste E, Walsh DA (2005). "Does the 'Ring of Life' ring true?". Trends Microbiol. 13 (6): 256–61. doi:10.1016/j.tim.2005.03.012. PMID 15936656.
- ↑ Doolittle WF, Bapteste E (February 2007). "Pattern pluralism and the Tree of Life hypothesis". Proc. Natl. Acad. Sci. U.S.A. 104 (7): 2043–9. doi:10.1073/pnas.0610699104. PMID 17261804. PMC 1892968. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=17261804.
- ↑ Kunin V, Goldovsky L, Darzentas N, Ouzounis CA (2005). "The net of life: reconstructing the microbial phylogenetic network". Genome Res. 15 (7): 954–9. doi:10.1101/gr.3666505. PMID 15965028. PMC 1172039. http://www.genome.org/cgi/pmidlookup?view=long&pmid=15965028.
- ↑ Jablonski D (1999). "The future of the fossil record". Science 284 (5423): 2114–16. doi:10.1126/science.284.5423.2114. PMID 10381868.
- ↑ Mason SF (1984). "Origins of biomolecular handedness". Nature 311 (5981): 19–23. doi:10.1038/311019a0. PMID 6472461.
- ↑ Wolf YI, Rogozin IB, Grishin NV, Koonin EV (2002). "Genome trees and the tree of life". Trends Genet. 18 (9): 472–79. doi:10.1016/S0168-9525(02)02744-0. PMID 12175808.
- ↑ Varki A, Altheide TK (2005). "Comparing the human and chimpanzee genomes: searching for needles in a haystack". Genome Res. 15 (12): 1746–58. doi:10.1101/gr.3737405. PMID 16339373.
- ↑ Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P (2006). "Toward automatic reconstruction of a highly resolved tree of life". Science 311 (5765): 1283–87. doi:10.1126/science.1123061. PMID 16513982.
- ↑ 240.0 240.1 Cavalier-Smith T (2006). "Cell evolution and Earth history: stasis and revolution". Philos Trans R Soc Lond B Biol Sci 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. PMID 16754610.
- ↑ Schopf J (2006). "Fossil evidence of Archaean life". Philos Trans R Soc Lond B Biol Sci 361 (1470): 869–85. doi:10.1098/rstb.2006.1834. PMID 16754604.
*Altermann W, Kazmierczak J (2003). "Archean microfossils: a reappraisal of early life on Earth". Res Microbiol 154 (9): 611–17. doi:10.1016/j.resmic.2003.08.006. PMID 14596897. - ↑ Schopf J (1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proc Natl Acad Sci U S a 91 (15): 6735–42. doi:10.1073/pnas.91.15.6735. PMID 8041691.
- ↑ Dyall S, Brown M, Johnson P (2004). "Ancient invasions: from endosymbionts to organelles". Science 304 (5668): 253–57. doi:10.1126/science.1094884. PMID 15073369.
- ↑ Martin W (2005). "The missing link between hydrogenosomes and mitochondria". Trends Microbiol. 13 (10): 457–59. doi:10.1016/j.tim.2005.08.005. PMID 16109488.
- ↑ Lang B, Gray M, Burger G (1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annu Rev Genet 33: 351–97. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
*McFadden G (1999). "Endosymbiosis and evolution of the plant cell". Curr Opin Plant Biol 2 (6): 513–19. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659. - ↑ DeLong E, Pace N (2001). "Environmental diversity of bacteria and archaea". Syst Biol 50 (4): 470–8. doi:10.1080/106351501750435040. PMID 12116647.
- ↑ Kaiser D (2001). "Building a multicellular organism". Annu. Rev. Genet. 35: 103–23. doi:10.1146/annurev.genet.35.102401.090145. PMID 11700279.
- ↑ Valentine JW, Jablonski D, Erwin DH (1 March 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Development 126 (5): 851–9. PMID 9927587. http://dev.biologists.org/cgi/reprint/126/5/851.
- ↑ Ohno S (1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". J. Mol. Evol. 44 Suppl 1: S23–7. doi:10.1007/PL00000055. PMID 9071008.
*Valentine J, Jablonski D (2003). "Morphological and developmental macroevolution: a paleontological perspective". Int. J. Dev. Biol. 47 (7–8): 517–22. PMID 14756327. http://www.ijdb.ehu.es/web/paper.php?doi=14756327. - ↑ Waters ER (2003). "Molecular adaptation and the origin of land plants". Mol. Phylogenet. Evol. 29 (3): 456–63. doi:10.1016/j.ympev.2003.07.018. PMID 14615186.
- ↑ Mayhew PJ (August 2007). "Why are there so many insect species? Perspectives from fossils and phylogenies". Biol Rev Camb Philos Soc 82 (3): 425–54. doi:10.1111/j.1469-185X.2007.00018.x. PMID 17624962.
- ↑ Bull JJ, Wichman HA (2001). "Applied evolution". Annu Rev Ecol Syst 32: 183–217. doi:10.1146/annurev.ecolsys.32.081501.114020.
- ↑ Doebley JF, Gaut BS, Smith BD (2006). "The molecular genetics of crop domestication". Cell 127 (7): 1309–21. doi:10.1016/j.cell.2006.12.006. PMID 17190597.
- ↑ Jäckel C, Kast P, Hilvert D (2008). "Protein design by directed evolution". Annu Rev Biophys 37: 153–73. doi:10.1146/annurev.biophys.37.032807.125832. PMID 18573077.
- ↑ Maher B. (April 2009). "Evolution: Biology's next top model?". Nature 458 (7239): 695–8. doi:10.1038/458695a. PMID 19360058.
- ↑ Borowsky R (January 2008). "Restoring sight in blind cavefish". Curr. Biol. 18 (1): R23–4. doi:10.1016/j.cub.2007.11.023. PMID 18177707.
- ↑ Gross JB, Borowsky R, Tabin CJ (January 2009). "A novel role for Mc1r in the parallel evolution of depigmentation in independent populations of the cavefish Astyanax mexicanus". PLoS Genet. 5 (1): e1000326. doi:10.1371/journal.pgen.1000326. PMID 19119422.
- ↑ Yergeau DA, Cornell CN, Parker SK, Zhou Y, Detrich HW (July 2005). "bloodthirsty, an RBCC/TRIM gene required for erythropoiesis in zebrafish". Dev. Biol. 283 (1): 97–112. doi:10.1016/j.ydbio.2005.04.006. PMID 15890331.
- ↑ Fraser AS (1958). "Monte Carlo analyses of genetic models". Nature 181 (4603): 208–9. doi:10.1038/181208a0. PMID 13504138.
- ↑ Rechenberg, Ingo (1973) (in German). Evolutionsstrategie – Optimierung technischer Systeme nach Prinzipien der biologischen Evolution (PhD thesis). Fromman-Holzboog.
- ↑ Holland, John H. (1975). Adaptation in Natural and Artificial Systems. University of Michigan Press. ISBN 0262581116.
- ↑ Koza, John R. (1992). Genetic Programming. MIT Press. ISBN 0262111705.
- ↑ Jamshidi M (2003). "Tools for intelligent control: fuzzy controllers, neural networks and genetic algorithms". Philosophical transactions. Series A, Mathematical, physical, and engineering sciences 361 (1809): 1781–808. doi:10.1098/rsta.2003.1225. PMID 12952685.
- ↑ Browne, Janet (2003). Charles Darwin: The Power of Place. London: Pimlico. pp. 376–379. ISBN 0-7126-6837-3.
- ↑ For an overview of the philosophical, religious, and cosmological controversies, see: Dennett, D (1995). Darwin's Dangerous Idea: Evolution and the Meanings of Life. Simon & Schuster. ISBN 978-0684824710.
*For the scientific and social reception of evolution in the 19th and early 20th centuries, see: Johnston, Ian C.. "History of Science: Origins of Evolutionary Theory". And Still We Evolve. Liberal Studies Department, Malaspina University College. http://records.viu.ca/~johnstoi/darwin/sect3.htm. Retrieved 2007-05-24.
*Bowler, PJ (2003). Evolution: The History of an Idea, Third Edition, Completely Revised and Expanded. University of California Press. ISBN 978-0520236936.
*Zuckerkandl E (2006). "Intelligent design and biological complexity". Gene 385: 2–18. doi:10.1016/j.gene.2006.03.025. PMID 17011142. - ↑ Ross, M.R. (2005). "Who Believes What? Clearing up Confusion over Intelligent Design and Young-Earth Creationism" (PDF). Journal of Geoscience Education 53 (3): 319. http://www.nagt.org/files/nagt/jge/abstracts/Ross_v53n3p319.pdf. Retrieved 2008-04-28.
- ↑ Hameed, Salman (2008-12-12). "Science and Religion: Bracing for Islamic Creationism". Science 322 (5908): 1637–1638. doi:10.1126/science.1163672. PMID 19074331. http://helios.hampshire.edu/~sahCS/Hameed-Science-Creationism.pdf. Retrieved 2009.
- ↑ Spergel D. N.; Scott, EC; Okamoto, S (2006). "Science communication. Public acceptance of evolution". Science 313 (5788): 765–66. doi:10.1126/science.1126746. PMID 16902112.
- ↑ Spergel, D. N.; Verde, L.; Peiris, H. V.; Komatsu, E.; Nolta, M. R.; Bennett, C. L.; Halpern, M.; Hinshaw, G. et al. (2003). "First-Year Wilkinson Microwave Anisotropy Probe (WMAP) Observations: Determination of Cosmological Parameters". The Astrophysical Journal Supplement Series 148: 175–94. doi:10.1086/377226.
- ↑ Wilde SA, Valley JW, Peck WH, Graham CM (2001). "Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago". Nature 409 (6817): 175–78. doi:10.1038/35051550. PMID 11196637.
- ↑ Understanding Creationism after Kitzmiller 2007
- ↑ On the history of eugenics and evolution, see Kevles, D (1998). In the Name of Eugenics: Genetics and the Uses of Human Heredity. Harvard University Press. ISBN 978-0674445574.
- ↑ Darwin strongly disagreed with attempts by Herbert Spencer and others to extrapolate evolutionary ideas to all possible subjects; see Midgley, M (2004). The Myths we Live By. Routledge. p. 62. ISBN 978-0415340779.
- ↑ Allhoff F (2003). "Evolutionary ethics from Darwin to Moore". History and philosophy of the life sciences 25 (1): 51–79. doi:10.1080/03919710312331272945. PMID 15293515.
- ↑ Gowaty, Patricia Adair (1997). Feminism and evolutionary biology: boundaries, intersections, and frontiers. London: Chapman & Hall. ISBN 0-412-07361-7.
Further reading
- Introductory reading
- Carroll, S. (2005). Endless Forms Most Beautiful. New York: W.W. Norton. ISBN 0-393-06016-0.
- Charlesworth, C.B. and Charlesworth, D. (2003). Evolution. Oxfordshire: Oxford University Press. ISBN 0-192-80251-8.
- Dawkins, R. (2006). The Selfish Gene: 30th Anniversary Edition. Oxford University Press. ISBN 0199291152.
- Gould, S.J. (1989). Wonderful Life: The Burgess Shale and the Nature of History. New York: W.W. Norton. ISBN 0-393-30700-X.
- Jones, S. (2001). Almost Like a Whale: The Origin of Species Updated. (American title: Darwin's Ghost). New York: Ballantine Books. ISBN 0-345-42277-5.
- Mader, Sylvia S. (2007). Biology. Murray P. Pendarvis (9th ed.). McGraw Hill. ISBN 9780073258393.
- Maynard Smith, J. (1993). The Theory of Evolution: Canto Edition. Cambridge University Press. ISBN 0-521-45128-0.
- Pallen, M.J. (2009). The Rough Guide to Evolution. Rough Guides. ISBN 978-1-85828-946-5.
- Smith, C.B. and Sullivan, C. (2007). The Top 10 Myths about Evolution. Prometheus Books. ISBN 978-1-59102-479-8.
- History of evolutionary thought
- Darwin, Charles (1859). On the Origin of Species (1st ed.). London: John Murray. ISBN 0801413192. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=1.
- Larson, E.J. (2004). Evolution: The Remarkable History of a Scientific Theory. New York: Modern Library. ISBN 0-679-64288-9.
- Zimmer, C. (2001). Evolution: The Triumph of an Idea. London: HarperCollins. ISBN 0-060-19906-7.
- Advanced reading
- Barton, N.H., Briggs, D.E.G., Eisen, J.A., Goldstein, D.B. and Patel, N.H. (2007). Evolution. Cold Spring Harbor Laboratory Press. ISBN 0-879-69684-2.
- Coyne, J.A. and Orr, H.A. (2004). Speciation. Sunderland: Sinauer Associates. ISBN 0-878-93089-2.
- Futuyma, D.J. (2005). Evolution. Sunderland: Sinauer Associates. ISBN 0-878-93187-2.
- Gould, S.J. (2002). The Structure of Evolutionary Theory. Cambridge: Belknap Press (Harvard University Press). ISBN 0-674-00613-5.
- Maynard Smith, J. and Szathmáry, E. (1997). The Major Transitions in Evolution. Oxfordshire: Oxford University Press. ISBN 0-198-50294-X.
- Mayr, E. (2001). What Evolution Is. New York: Basic Books. ISBN 0-465-04426-3.
- Olson, Wendy; Hall, Brian Keith (2003). Keywords and concepts in evolutionary developmental biology. Cambridge: Harvard University Press. ISBN 0-674-02240-8.
External links
- General information
- Everything you wanted to know about evolution by New Scientist
- Howstuffworks.com — How Evolution Works
- National Academies Evolution Resources
- Synthetic Theory Of Evolution: An Introduction to Modern Evolutionary Concepts and Theories
- Understanding Evolution from University of California, Berkeley
- Evolution of Evolution - 150 Years of Darwin's "On the Origin of Species"
- History of evolutionary thought
- The Complete Work of Charles Darwin Online
- Understanding Evolution: History, Theory, Evidence, and Implications
- On-line lectures
- What Genomes Can Tell Us About the Past – lecture by Sydney Brenner
- The Origin of Vertebrates – lecture by Marc Kirschner
- The Making of the Fittest – lecture by Sean B. Carroll
|
||||||||||||||||||||||||||||||||